引用本文: 支知娜, 孫翠敏, 傅芊, 陳斯, 周翔天. 小鼠形覺剝奪對不同類型視網膜神經節細胞形態的影響. 中華眼底病雜志, 2019, 35(5): 451-461. doi: 10.3760/cma.j.issn.1005-1015.2019.05.006 復制
屈光發育關鍵期,合適的視覺輸入是正視化的關鍵因子[1-2]。RGC是視網膜上將視覺信號傳遞到中樞的唯一神經元。不同的視覺信號特征由不同的視網膜神經通路及相應的RGC來承擔[3-5]。視覺信號可分為形覺信號和非形覺信號。形覺信號包括物體的形狀、運動等特征,由傳統的RGC傳遞;非形覺信號參與晝夜節律和瞳孔對光反射等視網膜功能,主要由感光RGC(ipRGC)進行傳遞[3-5]。形覺和非形覺視覺信號在近視的發生發展中均發揮了重要的作用。既往研究發現,超過一半的OFF瞬時性RGC和ON持續性RGC對遠視性和近視性離焦的反應有所不同[6];兔子形覺剝奪過程中RGC的數量減少[7]。這些研究結果證實,RGC功能和數量在近視發生的過程中發揮了作用。RGC的形態是其功能的基礎。比如,局部邊緣探測RGC的樹突野小,分支密集;而ipRGC樹突野大,分支疏松[8]。因此,RGC改變可以反應哪些RGC受形覺剝奪的影響而參與近視過程,以及不同類型的RGC及其所代表的通路是否在近視過程中發揮了不同作用值得深入研究。本研究觀察分析了黃色熒光蛋白(YFP)轉基因小鼠形覺剝奪對不同類型RGC形態的影響,以期為進一步明確近視的視網膜機制提供理論基礎。現將結果報道如下。
1 材料和方法
1.1 實驗動物及其基因型鑒定、實驗設計
本實驗所涉及動物飼養與處理均經溫州醫科大學動物倫理委員會審批通過,并遵循美國視覺與眼科學研究學會關于眼科和視覺研究中對動物的處理原則。60只B6.Cg-Tg(Thy1-YFP)HJrs/J轉基因小鼠(YFP轉基因小鼠)購自美國杰克遜實驗室,繁殖于溫州醫科大學動物實驗室(動物倫理批號WYDW2016-0348)。該小鼠特異性標記10%~30%的RGC。所有小鼠來自YFP轉基因小鼠同窩仔,均在12 h光照/12 h黑暗的照明條件下飼養于標準透光鼠籠中,光照時間為8:00至20:00,環境溫度(22±2)℃。可自由攝食、進水。每3天更換清潔籠具及墊料以保持飼養環境衛生。
剪取小鼠尾巴提取DNA,經PCR后電泳。YFP轉基因小鼠由C57BL/6J為背景,利用轉基因細菌人工染色體技術獲得。在表達胸腺細胞抗原1(Thy1)的基因片段上加上表達YFP的基因片段,雜合子小鼠體內所有表達Thy1的細胞均表達YFP。YFP陽性小鼠Thy1基因片段長度為415 bp,YFP陰性小鼠Thy1基因片段長度為324 bp。
將60只小鼠隨機分為形覺剝奪組及對照組,分別為28、32只。形覺剝奪組小鼠右眼通過遮蓋片進行單眼遮蓋2周作為剝奪眼,同時納入對側眼進行分析。對照組小鼠不作任何處理,取右眼作為對照眼。形覺剝奪前及形覺剝奪后2周,測量兩組小鼠屈光度及眼軸相關參數;采用免疫組織化學染色對視網膜上表達YFP的RGC進行三維重構和形態分析;采用免疫熒光染色對RGC數量進行統計。
1.2 實驗方法
采用自行搭建的紅外偏心攝影驗光儀檢測兩組小鼠的屈光度,每只眼連續測量3次,取平均值。采用Cinescan A/B超聲診斷儀(法國Quantel Medical公司)中的A超探頭進行測量,檢測參數包括前房深度、晶狀體厚度、玻璃體腔深度和眼軸長度。測量前2~3 min用0.5%鹽酸丙美卡因滴眼液進行表面麻醉,每只眼重復測量10次,標準差<0.04 mm,取平均值。
小鼠經5%水合氯醛麻醉后取眼球,分離視網膜,并將其修剪成四葉草形狀。先將視網膜在PBS中清洗3次(10 min/次),然后置于含3% H2O2的PBS中室溫振蕩孵育20 min以去除內源性的過氧化物酶,最后PBS清洗3次(10 min/次)。加入封閉液(5%驢血清+1%牛血清白蛋白+0.2%賴氨酸+1 mol PBS)室溫下封閉2 h后,加入用封閉液稀釋的一抗(抗綠色熒光蛋白,1∶500,美國Novus Biologicals公司)4 ℃孵育2 d后加入用封閉液稀釋的二抗(生物素Biotin標記親和純化驢抗羊IgG二抗,1∶500,美國Jackson Immuno Research公司),室溫孵育2 h,PBS清洗3次(10 min/次)。然后將視網膜置入親和素-生物素復合體(1∶500)4 ℃過夜。PBS清洗后,再經0.1 mol/L三羥甲基氨基甲烷鹽酸鹽(Tris-HCl)清洗5 min。將視網膜置入DAB染色液中避光靜置數分鐘至切片顯色。最后0.1 mol/L Tris-HCl清洗10 min,PBS清洗2次(10 min/次),封片。
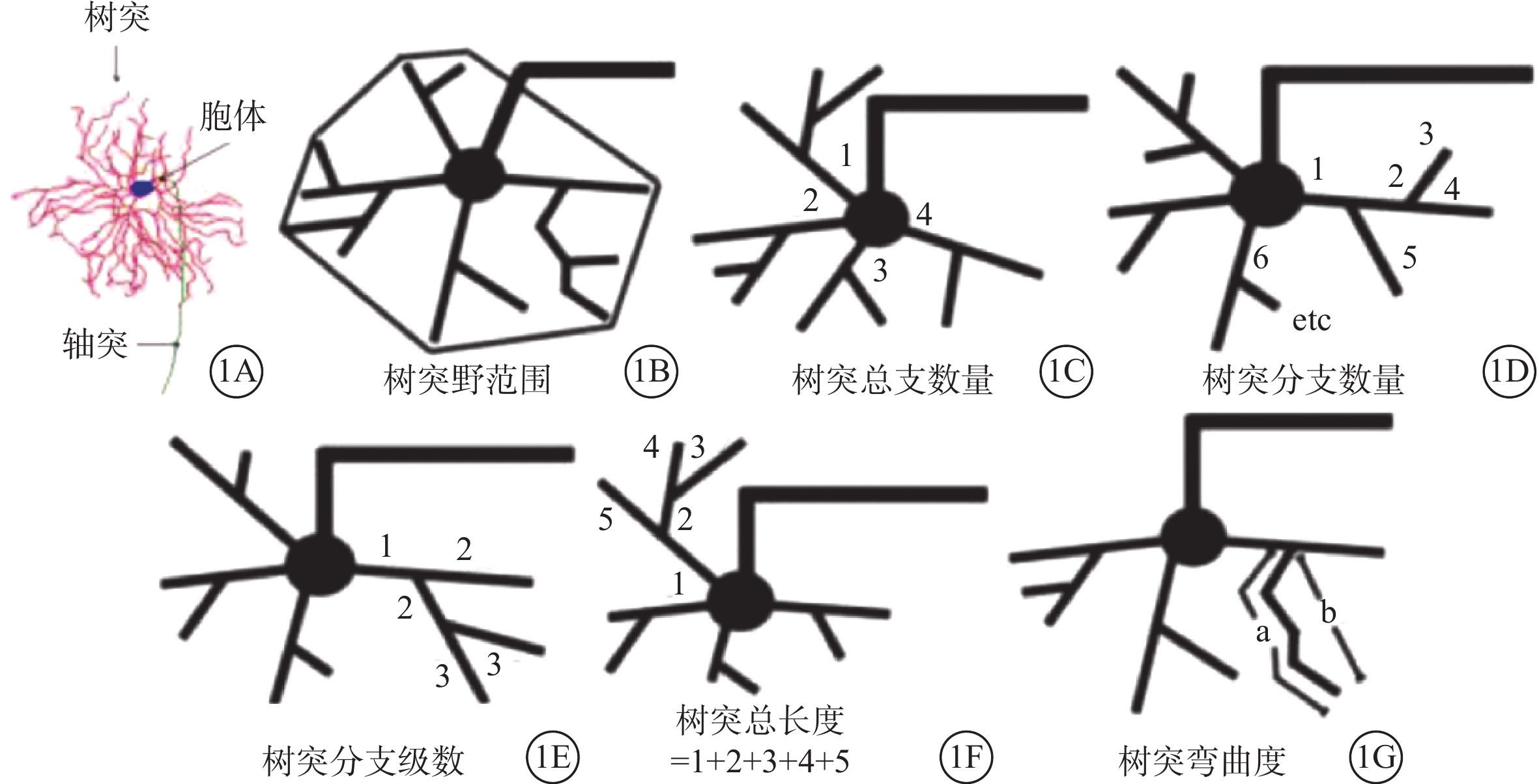
封片后的載玻片靜置晾干。在明場10倍鏡下,挑選顯色清晰、結構完整的RGC,先描繪視網膜的外輪廓。在63倍水鏡下,使用Neurolucida分析系統在線神經元三維重構系統重構目標神經元的三維立體結構,包括胞體、樹突和軸突(圖1A)。利用Neurolucida Explorer分析軟件對重構的RGC三維結構進行定量分析對比,包括胞體的周長及面積、樹突數量、樹突長度、樹突表面積、樹突體積、樹突的水平及垂直延展距離、樹突分級數、彎曲度及分支角度;同心圓分析獲得樹突分支點數目及分支長度等參數在內的23個形態學參數(圖1B~1G)。
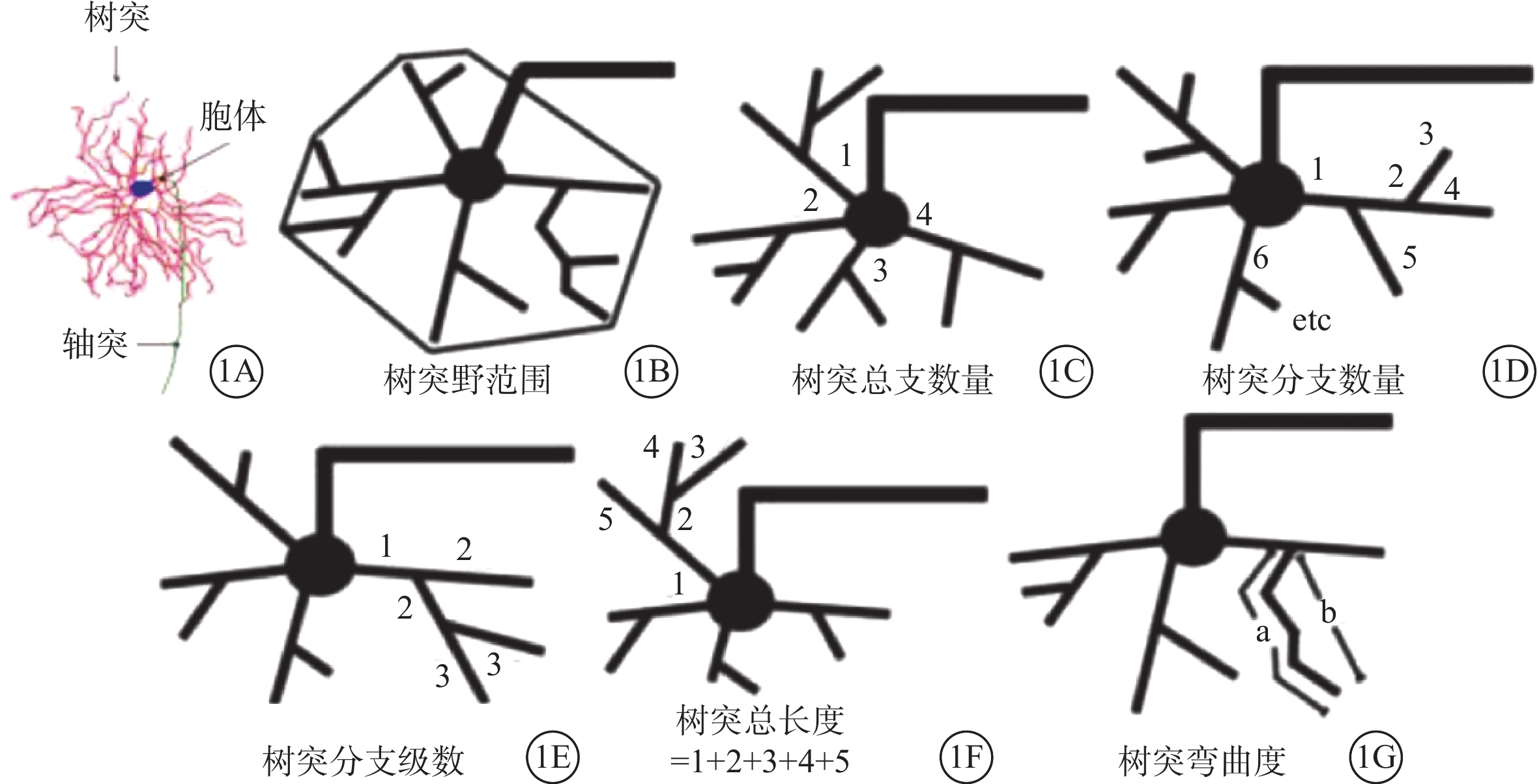 圖1
部分形態學參數分析示意圖
圖1
部分形態學參數分析示意圖
小鼠頸椎脫臼后取眼球,4%多聚甲醛溶液固定20 min后取視網膜并修剪成四葉草形狀。視網膜經0.1 mol PBS中清洗后加入封閉液(6%驢血清+1%牛血清白蛋白+ 0.2% Triton X-100,pH=7.4)室溫封閉2 h。然后加入一抗(Brn3a抗體,sc-31984,1:50,美國Santa Cruz公司),4 ℃孵育2 d,加入二抗(生物素Biotin標記親和純化驢抗羊IgG二抗,1:400,美國Jackson immuno Research公司)孵育2 h。PBS清洗3次(10 min/次),封片。利用共聚焦顯微鏡(Imager-Z1,德國Zeiss公司)在視網膜四葉草的近端和遠端兩個視野拍照,共取8個視野進行分析。計數DAPI標記的所有RCG層細胞和Brn3a抗體標記的RGC。
1.3 統計學方法
采用SPSS19.0軟件進行統計分析,數據用均數±標準差( )表示。形覺剝奪組剝奪眼及其對側眼、對照眼之間各參數比較采用單因素方差分析,并用最小顯著差法校正。組內左右眼之間各參數比較采用配對 t 檢驗。不同類型RGC占所有重構RGC的比例比較采用 χ2 檢驗。P<0.05為差異有統計學意義。
)表示。形覺剝奪組剝奪眼及其對側眼、對照眼之間各參數比較采用單因素方差分析,并用最小顯著差法校正。組內左右眼之間各參數比較采用配對 t 檢驗。不同類型RGC占所有重構RGC的比例比較采用 χ2 檢驗。P<0.05為差異有統計學意義。
2 結果
2.1 形覺剝奪對屈光度、眼軸相關參數及RGC數量的影響
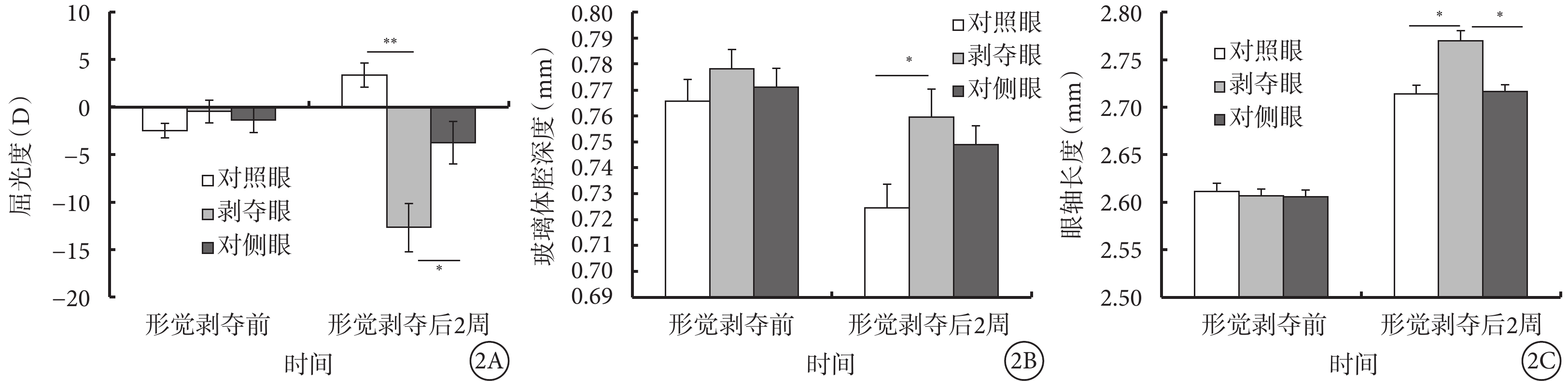
形覺剝奪前,形覺剝奪組剝奪眼、對側眼以及對照眼屈光度(F=0.849,P=0.436)和前房深度(F=0.197,P=0.822)、玻璃體腔深度(F=0.470,P=0.629)、眼軸長度(F=0.024,P=0.102)等眼軸相關參數比較,差異無統計學意義。形覺剝奪后2周,形覺剝奪組剝奪眼較對照眼出現了顯著的近視改變(F=15.009,P<0.001),同時伴有玻璃體腔深度加深(F=3.360,P=0.047)、眼軸延長(F=5.011,P=0.013);但前房深度和晶狀體厚度無明顯變化(P>0.05)(表1,圖2)。
 )
)
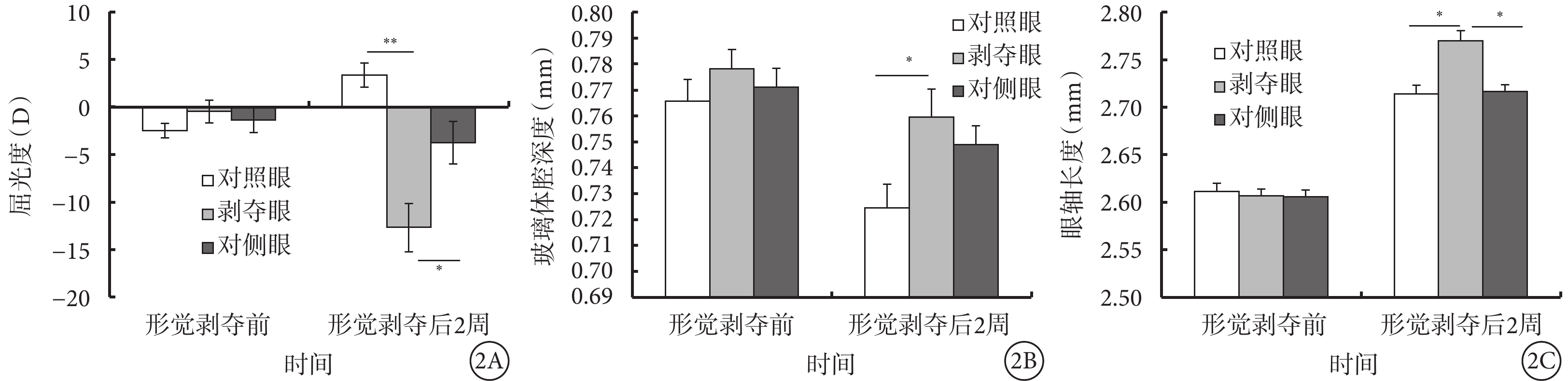 圖2
形覺剝奪后2周形覺剝奪組剝奪眼、對側眼及對照眼屈光度、眼軸相關參數比較。2A示屈光度;2B示玻璃體腔深度;2C示眼軸長度。*P<0.05,**P<0.001
圖2
形覺剝奪后2周形覺剝奪組剝奪眼、對側眼及對照眼屈光度、眼軸相關參數比較。2A示屈光度;2B示玻璃體腔深度;2C示眼軸長度。*P<0.05,**P<0.001
形覺剝奪組剝奪眼(n=4)、對側眼(n=3)以及對照眼(n=4)視網膜后極部RGC數量分別為(324±23)、(279±15)、(377±23)個,視網膜周邊部RGC數量分別為(270±31)、(288±8)、(310±47)個。在視網膜后極部,形覺剝奪組剝奪眼RGC數量較對側眼及對照組明顯減少,差異有統計學意義(F=4.769,P=0.035);但在周邊區,差異無統計學意義(F=1.517,P=0.266)。
2.2 RGC的類型及形態
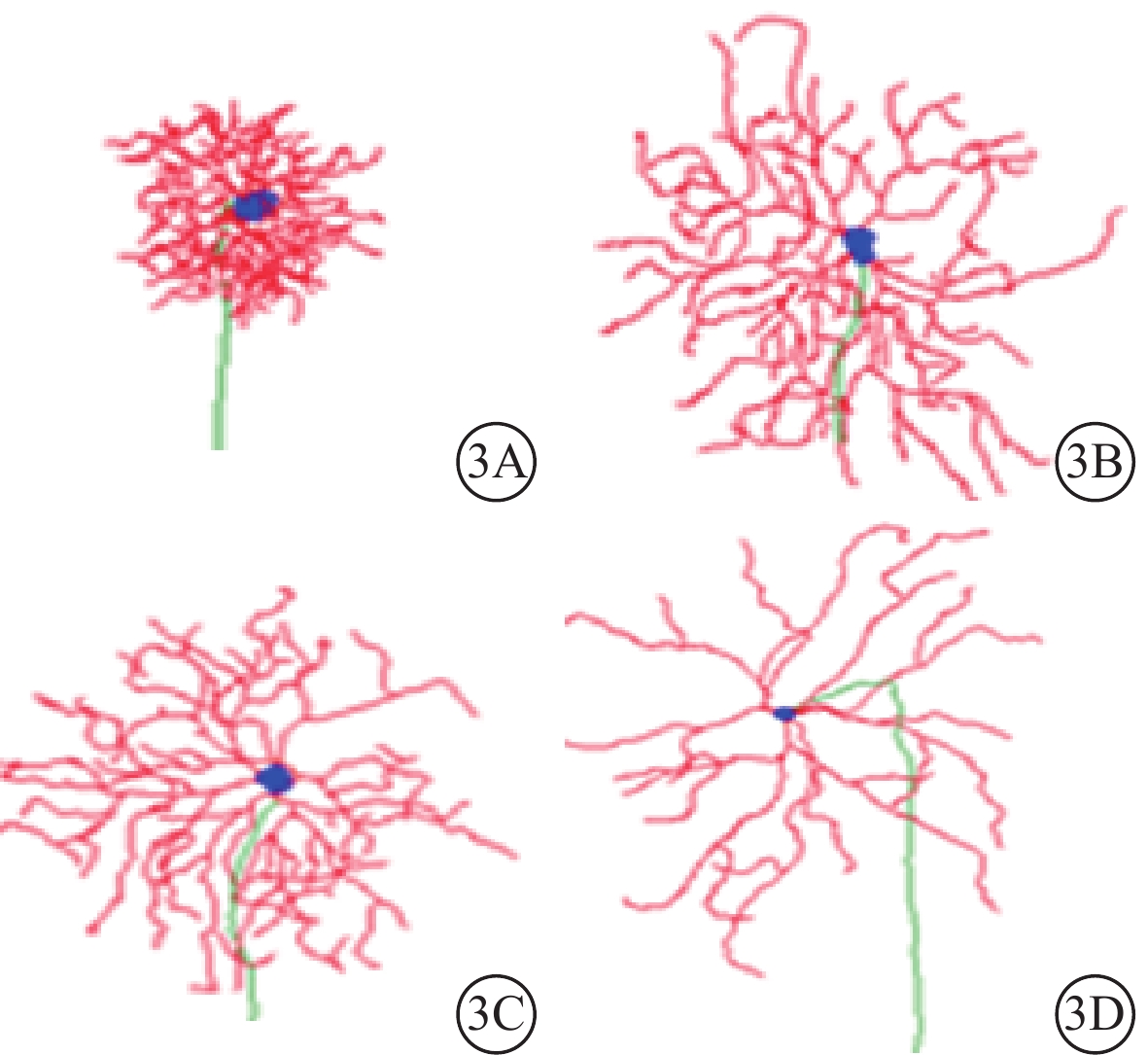
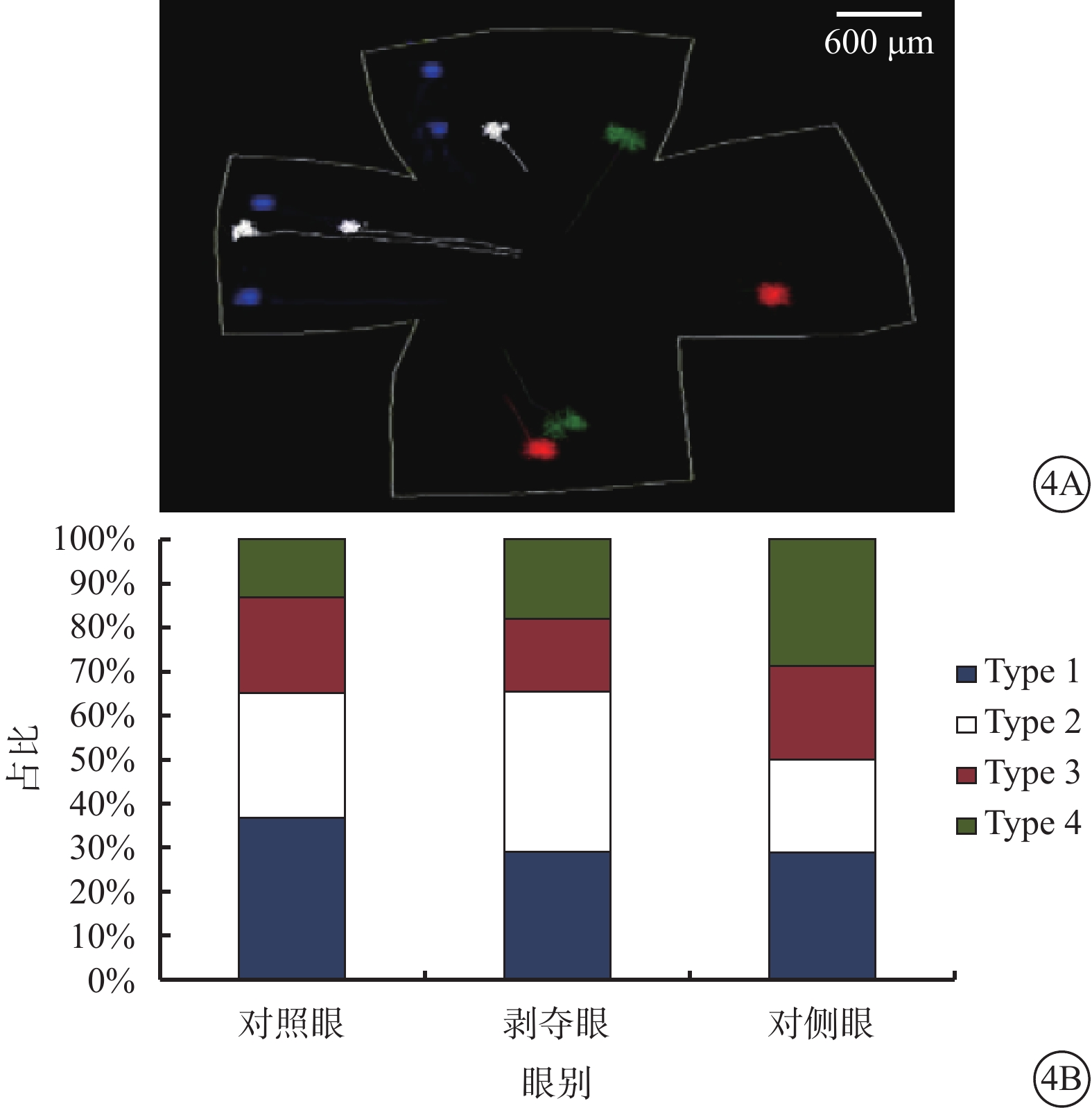
根據樹突野形態,三維重構的RGC可分為Type 1、Type 2、Type 3、Type 4共4種類型(圖3)。4種類型的RGC占比無明顯差異(χ2=6.853,P=0.335,各類型調整殘差≤2)(圖4)。
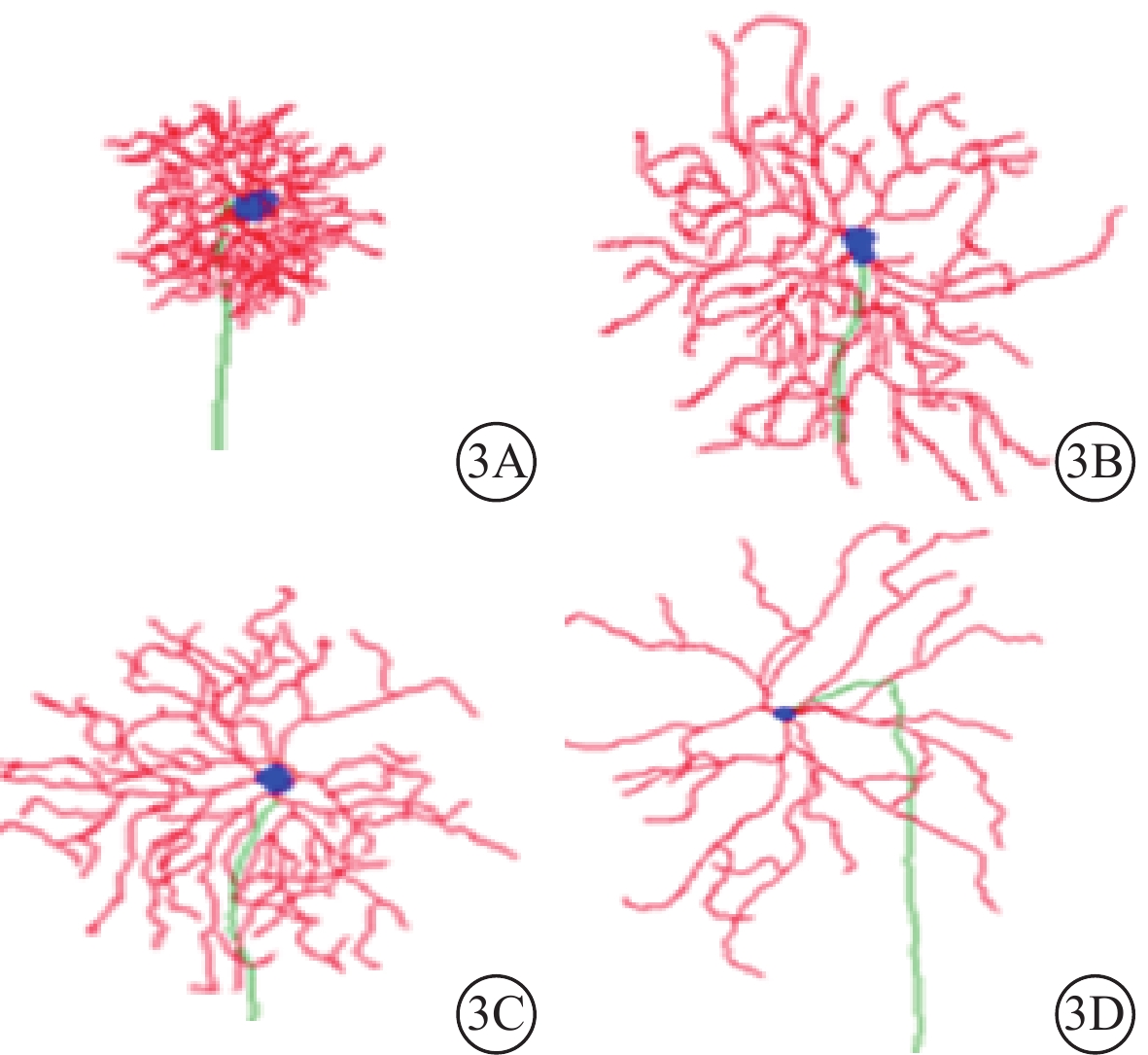 圖3
不同類型RGC形態示意圖。3A示Type 1;3B示Type 2;3C示Type 3;3D示Type 4
圖3
不同類型RGC形態示意圖。3A示Type 1;3B示Type 2;3C示Type 3;3D示Type 4
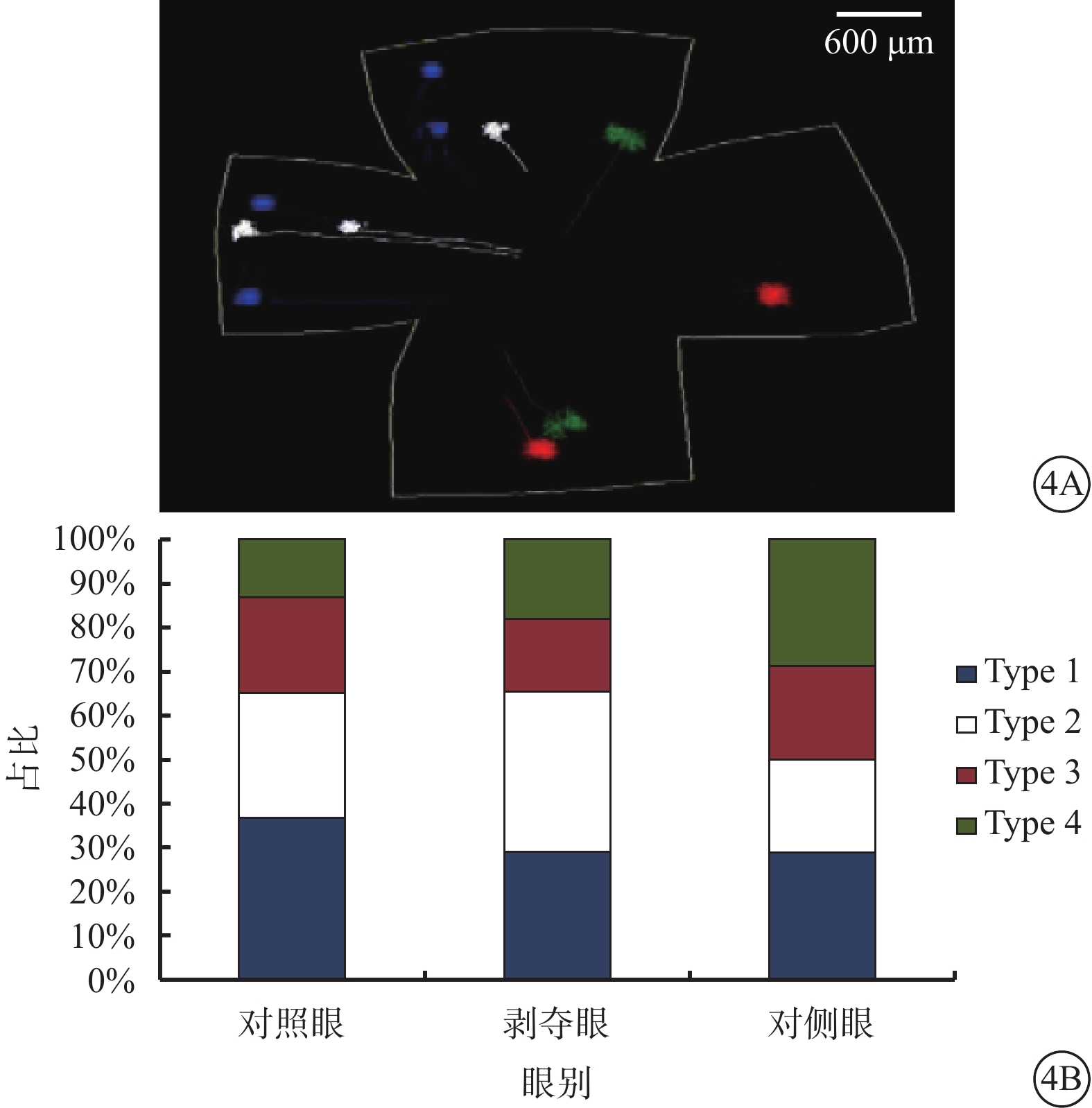 圖4
不同類型RGC在視網膜上的分布及其占各組重構細胞的比例。4A示4種類型RGC在視網膜上的分布示意圖,藍色代表Type 1,白色代表Type 2,紅色代表Type 3,綠色代表Type 4;4B示4種類型RGC占各組重構細胞的比例
圖4
不同類型RGC在視網膜上的分布及其占各組重構細胞的比例。4A示4種類型RGC在視網膜上的分布示意圖,藍色代表Type 1,白色代表Type 2,紅色代表Type 3,綠色代表Type 4;4B示4種類型RGC占各組重構細胞的比例
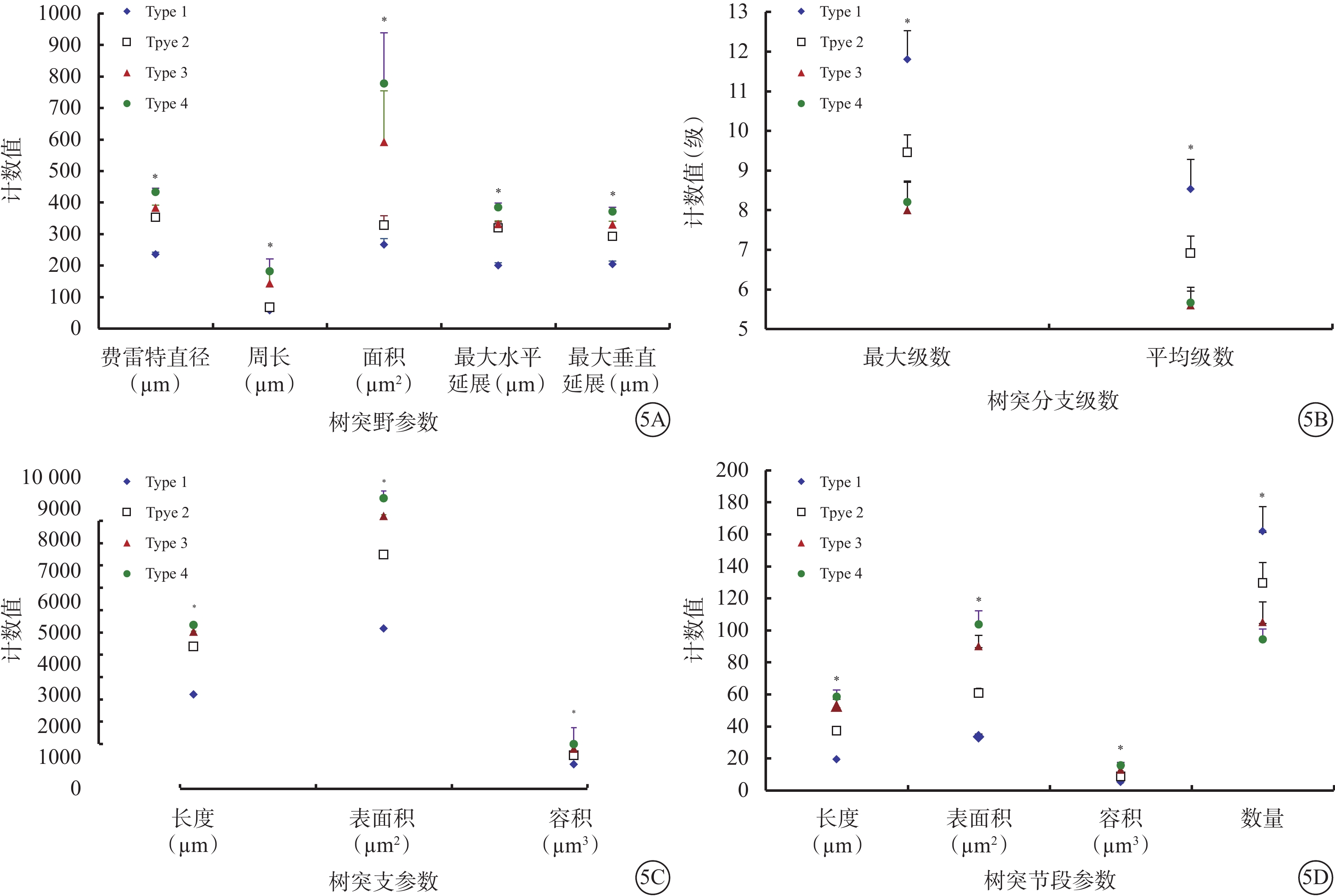
Type 1 RGC具有小而緊湊的樹突野;樹突支較少且短;樹突支分支級數、樹突節段多但短。與Type 1 RGC比較,Type 2 RGC樹突野更大;樹突分布較稀疏;樹突支數量更多且更長;樹突支分支級數、樹突節段數較少,但長度更長。與Type 2 RGC比較,Type 3 RGC樹突野更大,形態相似;樹突分布更豐富;樹突支數量更多且更長;樹突支分支級數、樹突節段數較少,但長度更長。Type 4 RGC樹突野形態是4種類型RGC中最大的;樹突分布最為稀疏;樹突支數量與Type 2、3 RGC相仿,長度略長;樹突支分支級數與Type 3 RGC接近;樹突節段數量與Type 2、3 RGC接近,但長度更長(表2,圖5)。
 )
)
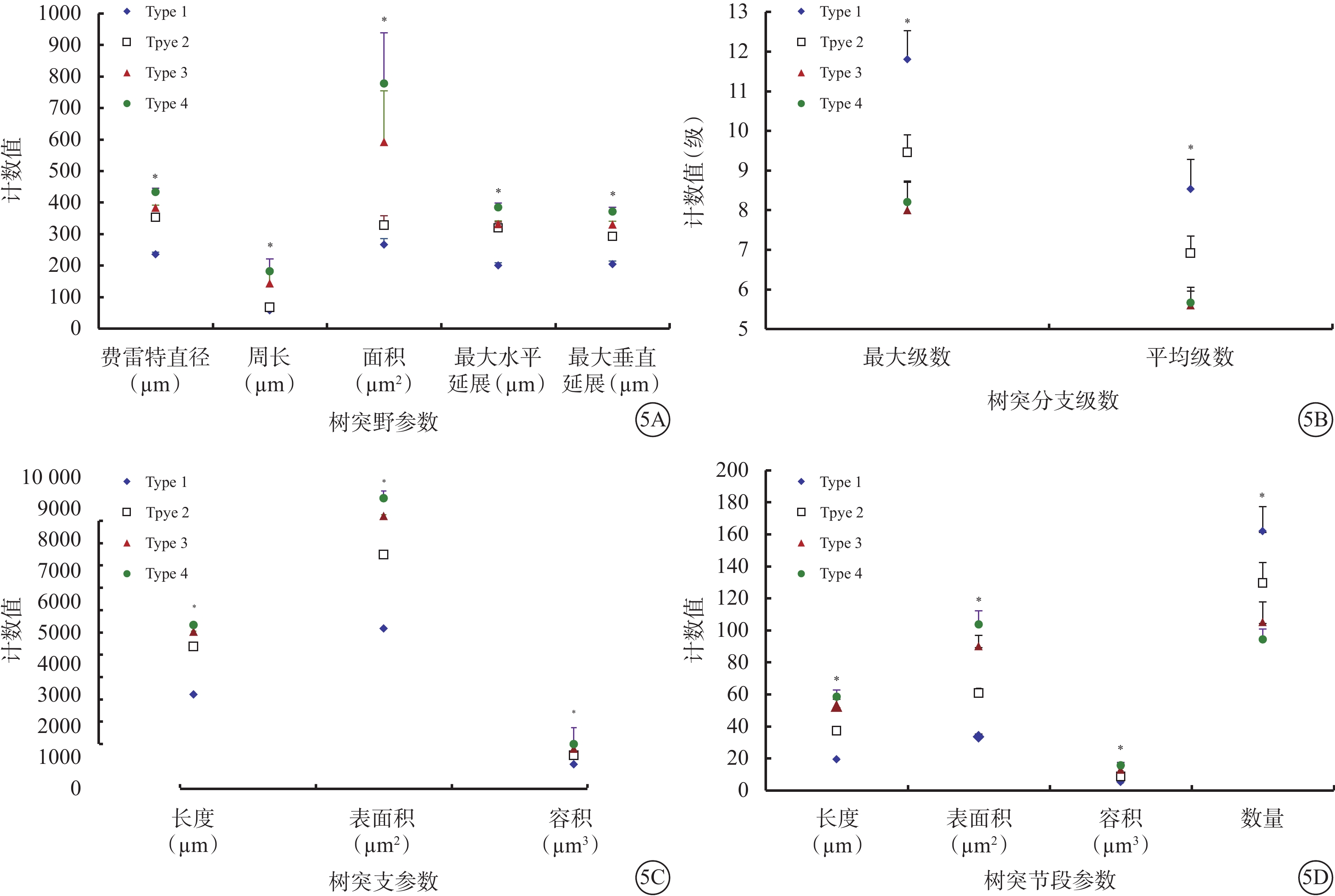 圖5
4種類型RGC樹突野、樹突支、樹突分支以及樹突節段等參數比較。5A示樹突野參數;5B示樹突支參數;5C示樹突分支參數;5D示樹突節段參數。*P<0.05
圖5
4種類型RGC樹突野、樹突支、樹突分支以及樹突節段等參數比較。5A示樹突野參數;5B示樹突支參數;5C示樹突分支參數;5D示樹突節段參數。*P<0.05
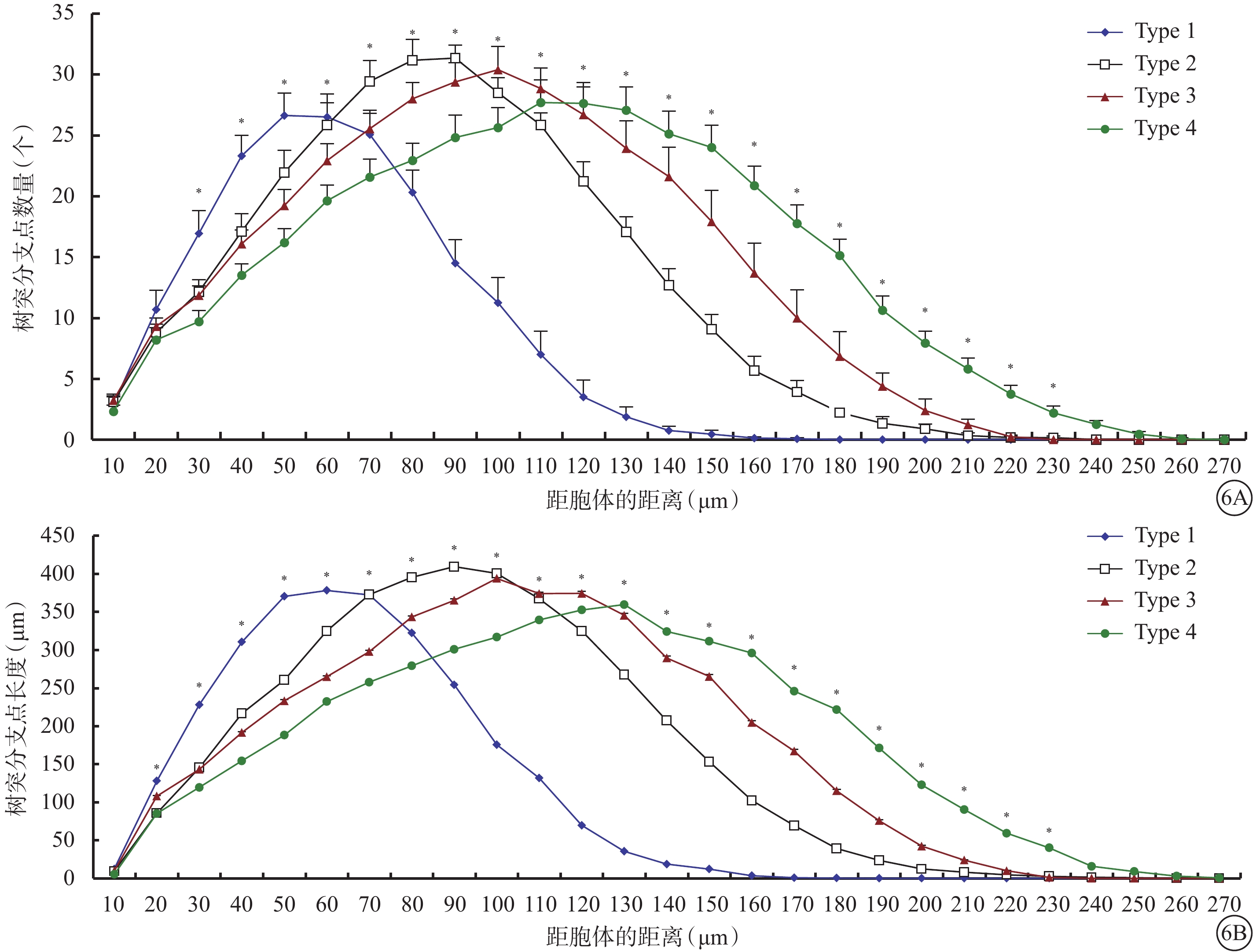
同心圓分析結果顯示,Type 1、Type 2、Type 3、Type 4 RGC樹突分支最多的區域分別集中在距離胞體50、90、100、130 μm處,分別在距離胞體140、200、210、240 μm處基本不再發出新的分支(圖6)。Type 3 RGC樹突分支最多的區域與Type 2 RGC接近,但在距離胞體110 μm以內樹突分支數量和長度較Type 2 RGC明顯少和短,在110 μm以外則樹突分支數量和長度均較Type 2 RGC更多。
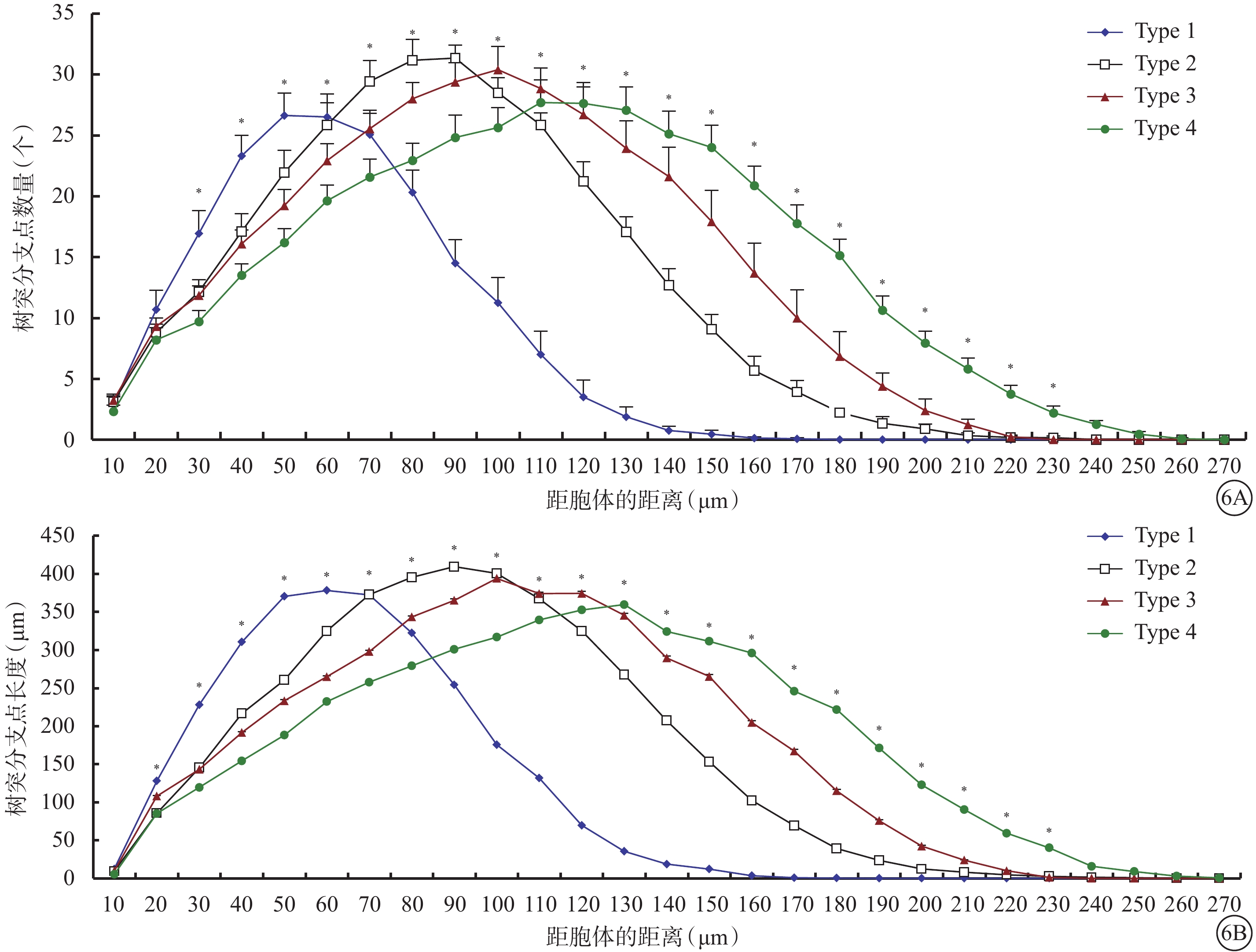 圖6
4種類型RGC同心圓分析圖。6A示離胞體不同距離內樹突分支點的數量;6B示離胞體不同距離內樹突分支長度。*P<0.05
圖6
4種類型RGC同心圓分析圖。6A示離胞體不同距離內樹突分支點的數量;6B示離胞體不同距離內樹突分支長度。*P<0.05
2.3 形覺剝奪對4種類型RGC形態的影響
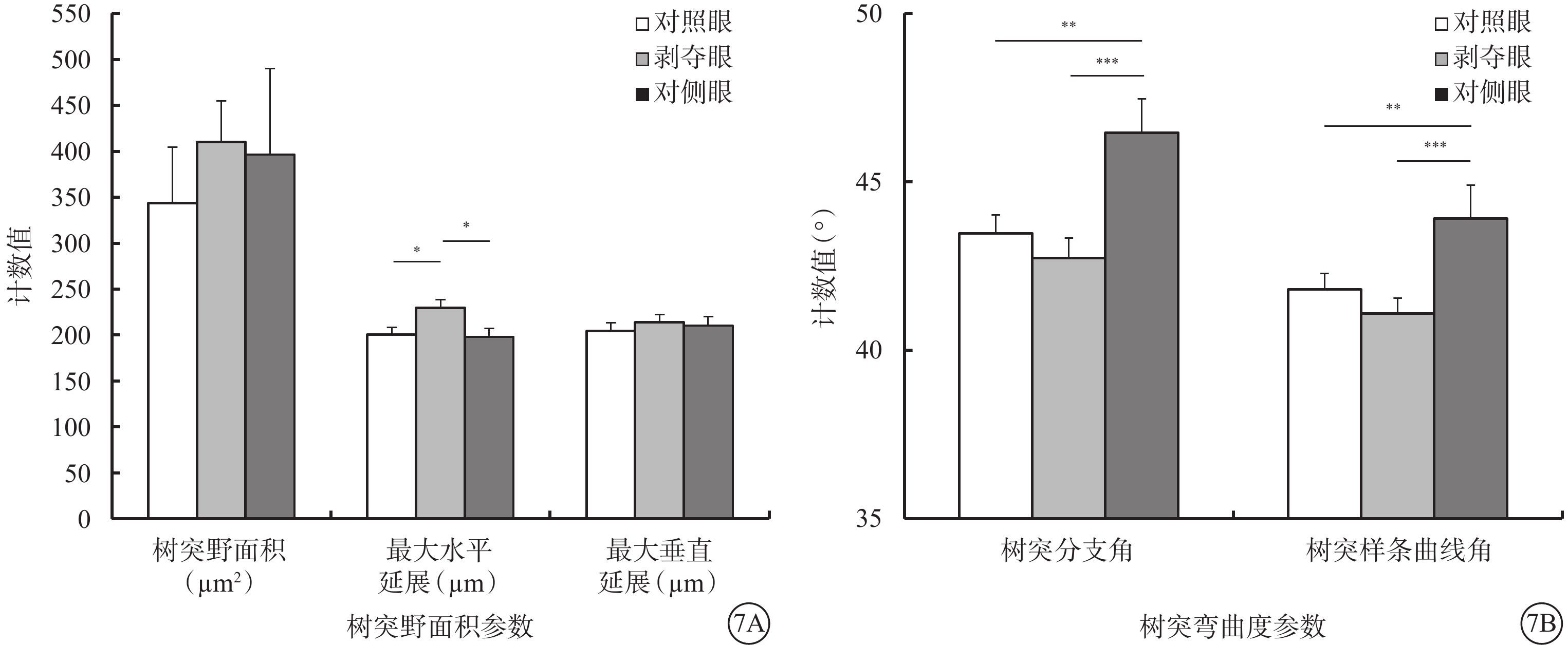
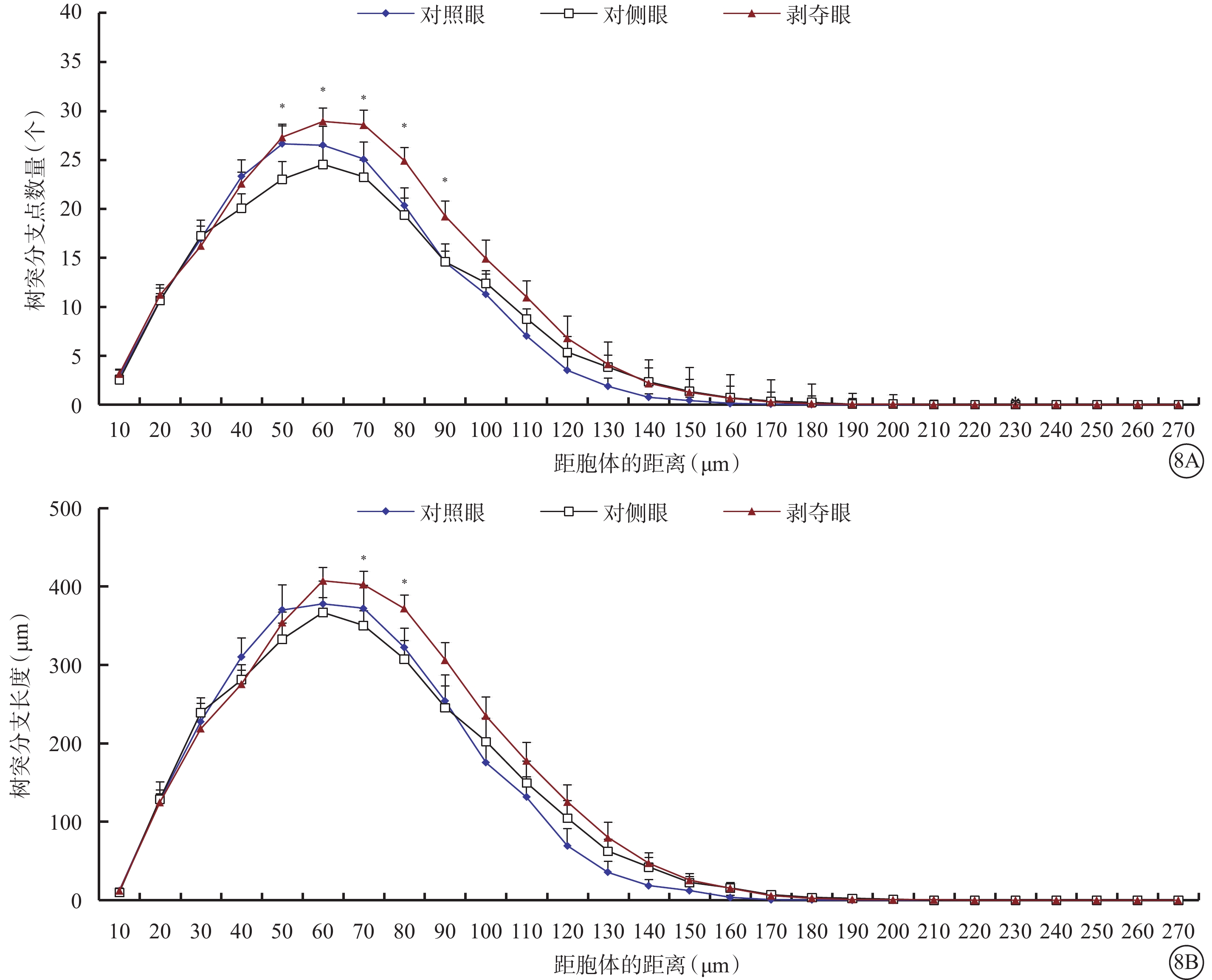
形覺剝奪對Type 1 RGC樹突野大小影響不大,但明顯增加了樹突野最大水平延展,降低了樹突彎曲度。與對側眼及對照眼比較,剝奪眼Type 1 RGC樹突野最大水平延展明顯增加(F=4.312,P=0.019),樹突彎曲度明顯下降(F=6.608,P=0.003),差異均有統計學意義(圖7)。同心圓分析結果顯示,形覺剝奪明顯增加了Type 1 RGC距離胞體50~90 μm區域內的樹突分支點數量,增長了樹突分支長度(圖8)。
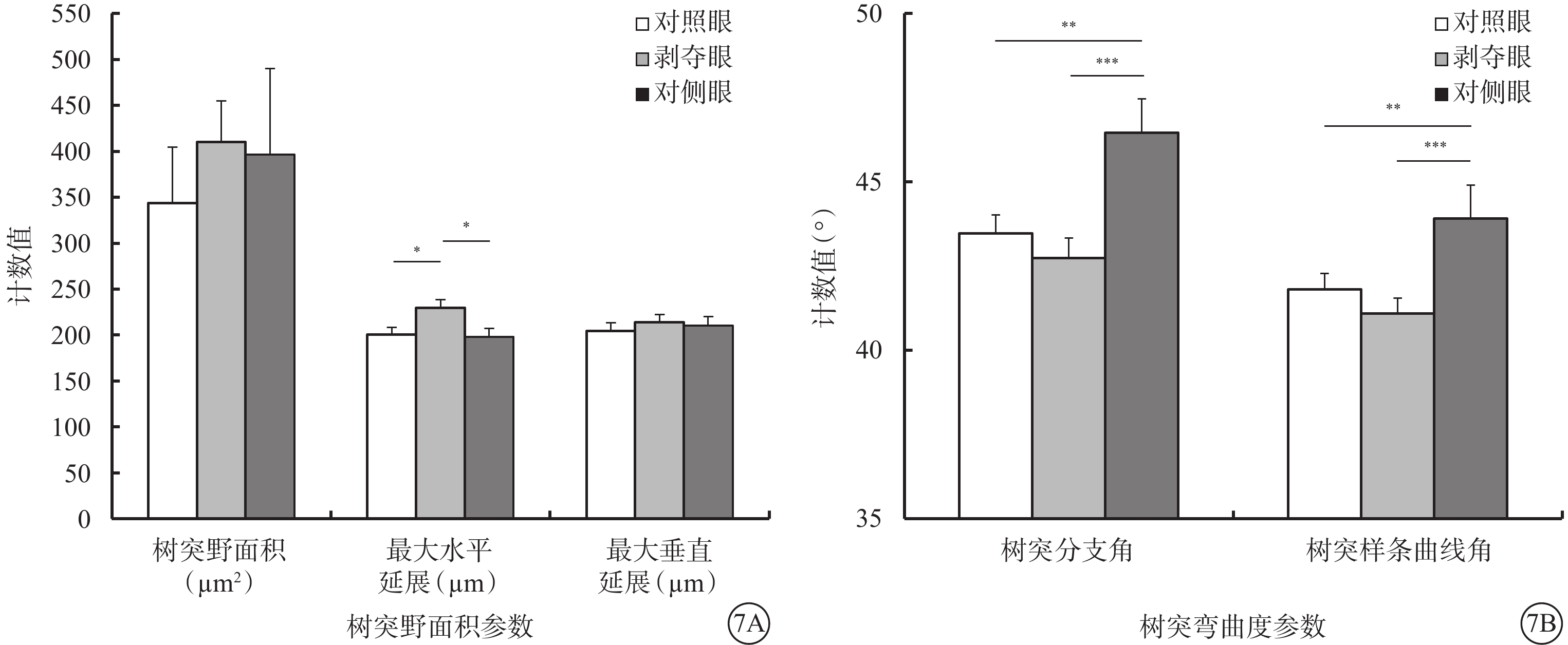 圖7
剝奪眼、對側眼及對照眼Type 1 RGC樹突野面積及樹突彎曲度參數比較。7A示樹突野面積參數;7B示樹突彎曲度參數。*P<0.05,**P<0.01,***P<0.001
圖7
剝奪眼、對側眼及對照眼Type 1 RGC樹突野面積及樹突彎曲度參數比較。7A示樹突野面積參數;7B示樹突彎曲度參數。*P<0.05,**P<0.01,***P<0.001
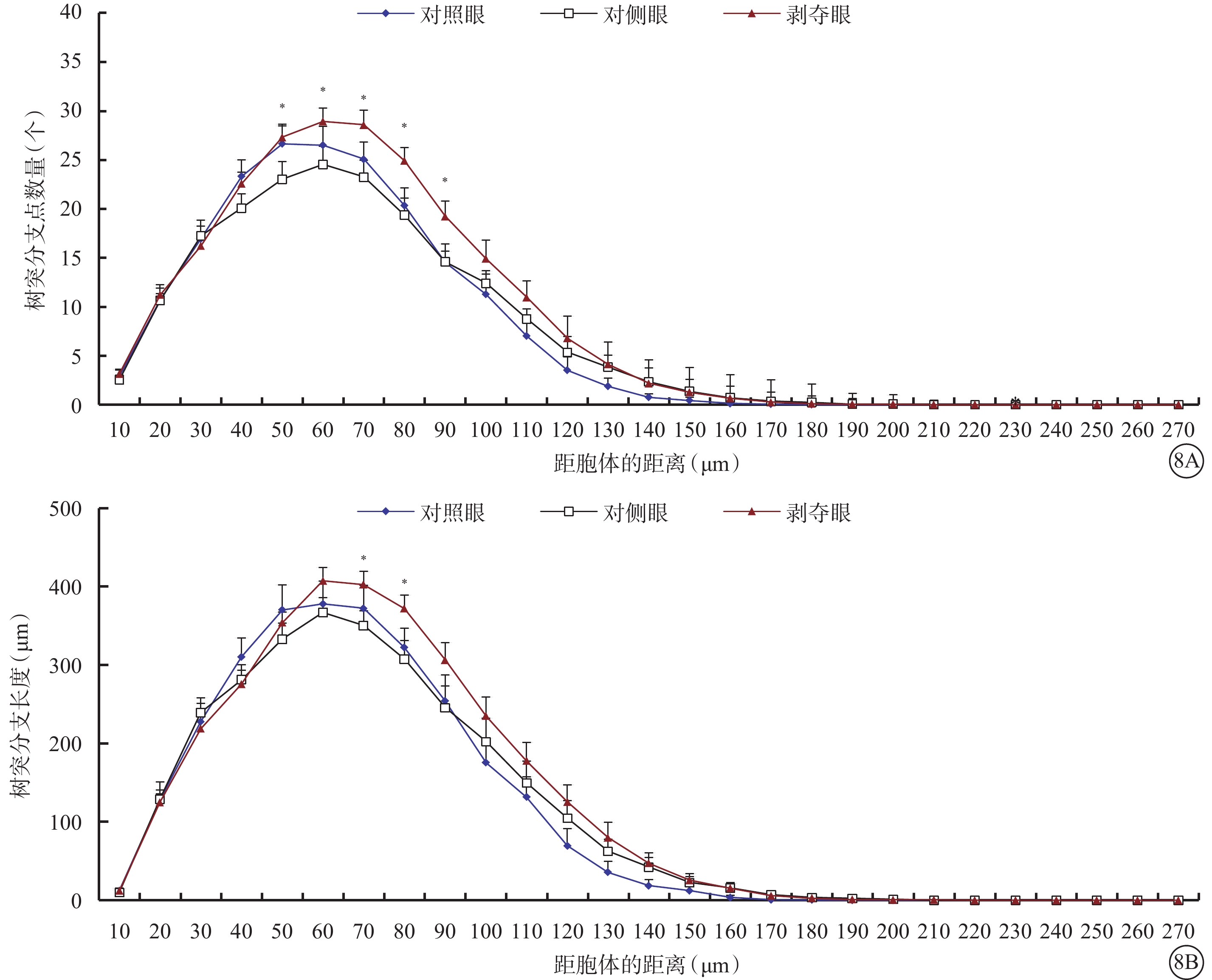 圖8
剝奪眼、對側眼及對照眼Type 1 RGC樹突分布同心圓分析圖。8A示剝奪眼Type 1 RGC距離胞體50~90 μm區域內的樹突分支點數量增多;8B示剝奪眼Type 1 RGC距離胞體50~90 μm區域內樹突分支長度增長。*P<0.05
圖8
剝奪眼、對側眼及對照眼Type 1 RGC樹突分布同心圓分析圖。8A示剝奪眼Type 1 RGC距離胞體50~90 μm區域內的樹突分支點數量增多;8B示剝奪眼Type 1 RGC距離胞體50~90 μm區域內樹突分支長度增長。*P<0.05
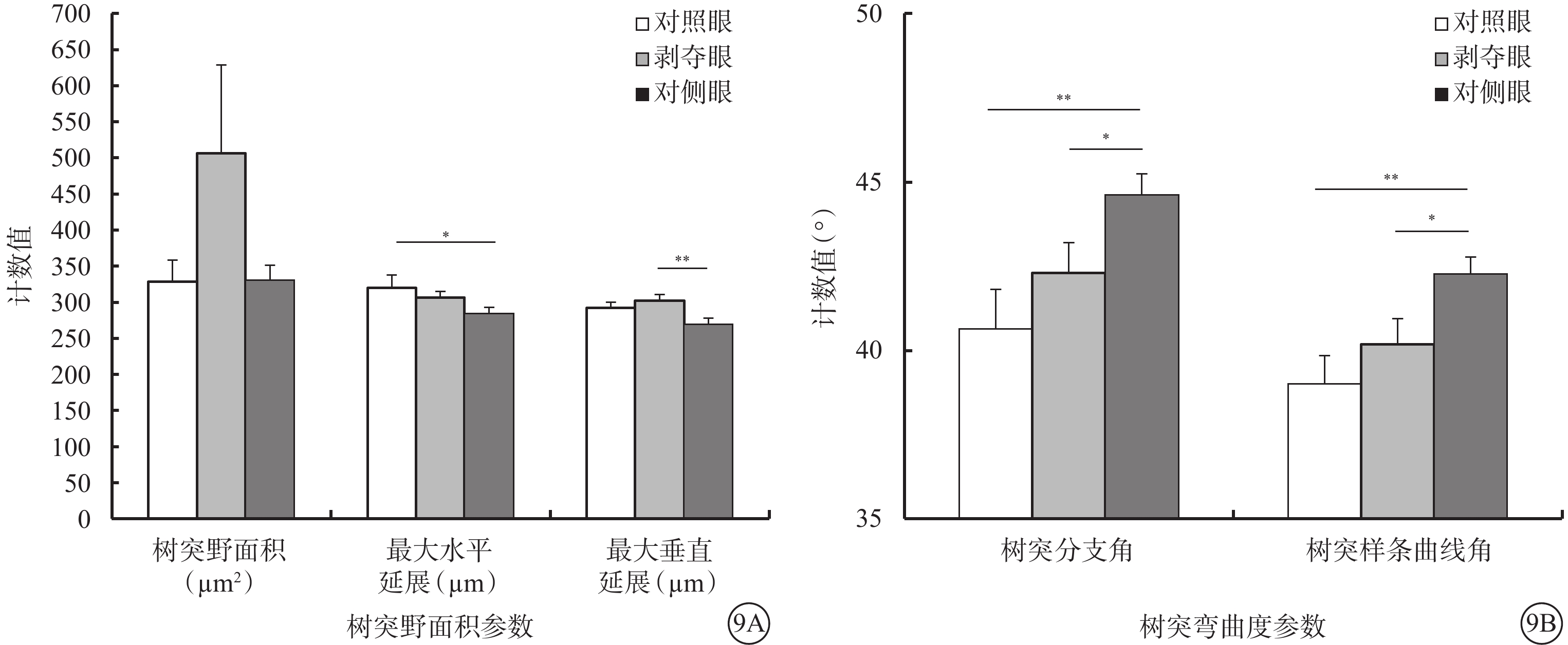
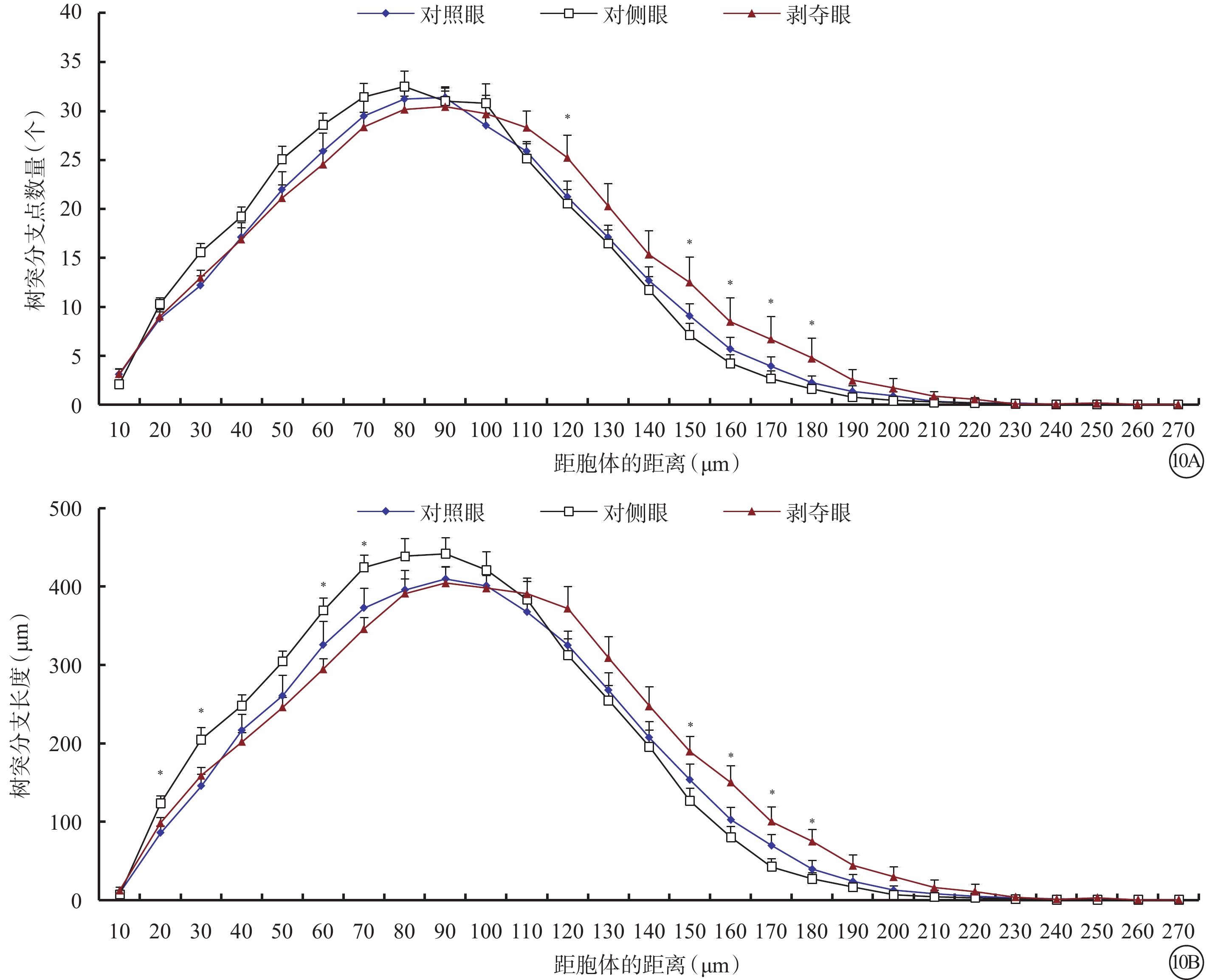
和Type1 RGC接近,形覺剝奪對Type 2 RGC的樹突野面積(F=1.780,P=0.180)以及最大水平延展(F=2.722,P=0.077)沒有明顯影響,但增加了其最大垂直延展(F=4.704,P=0.014)。同時,形覺剝奪減小了樹突分支角(F=5.180,P=0.009)以及樹突樣條曲線角度數(F=5.672,P=0.006)。剝奪眼Type 2 RGC樹突野最大垂直延展較對側眼明顯增加(P=0.012),但與對照眼無明顯差異(P=0.728)。剝奪眼Type 2 RGC樹突彎曲度和對側眼及對照眼相比均無明顯改變(F=2.606,P=0.085);但樹突分支角(P=0.046)、樹突樣條曲線角(P=0.026)較對側眼明顯變小(圖9)。同心圓分析結果顯示,形覺剝奪明顯縮短了20~30 μm及60~70 μm區域內Type 2 RGC樹突分支長度;增加了120~180 μm區域內Type 2 RGC樹突分支點數量以及150~180 μm區域內樹突分支長度(圖10)。
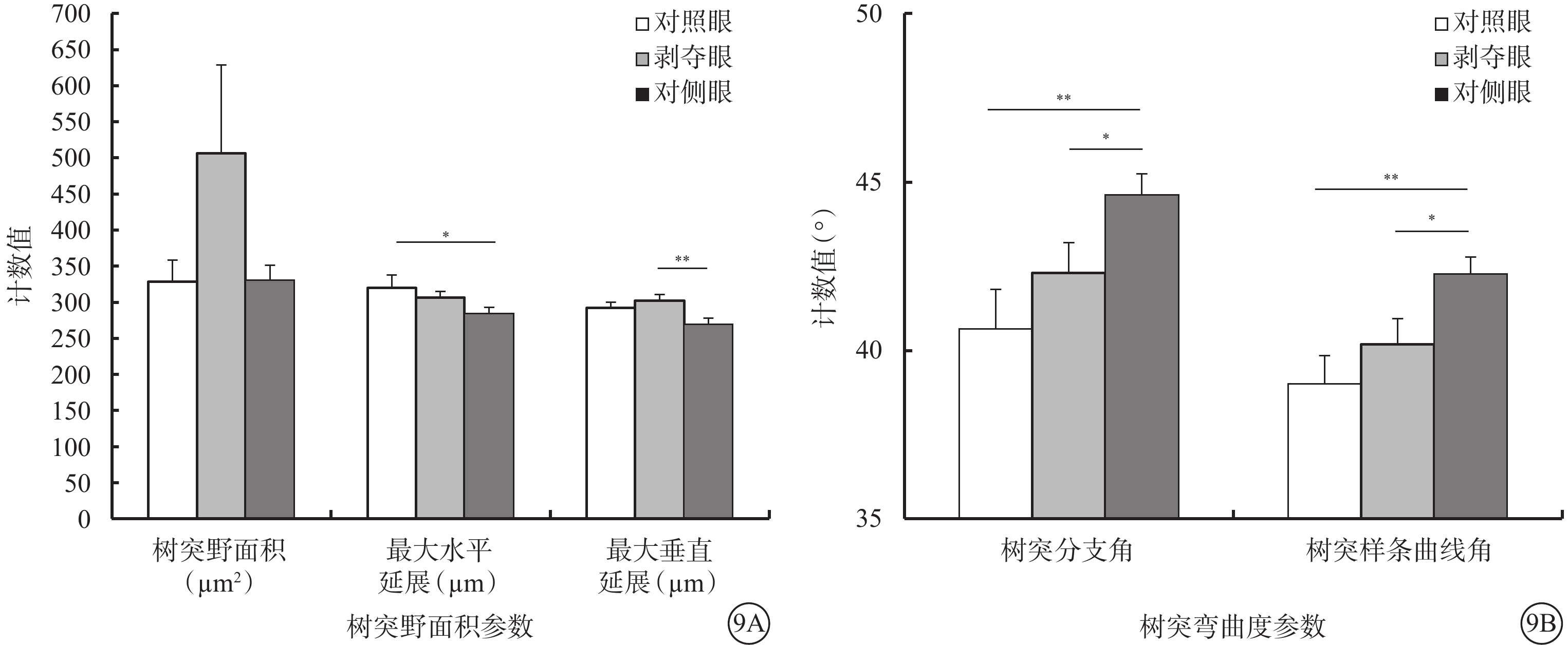 圖9
剝奪眼、對側眼及對照眼Type 2 RGC樹突野面積及樹突彎曲度參數比較。9A示樹突野面積參數;9B示樹突彎曲度參數。*P<0.05,**P<0.01
圖9
剝奪眼、對側眼及對照眼Type 2 RGC樹突野面積及樹突彎曲度參數比較。9A示樹突野面積參數;9B示樹突彎曲度參數。*P<0.05,**P<0.01
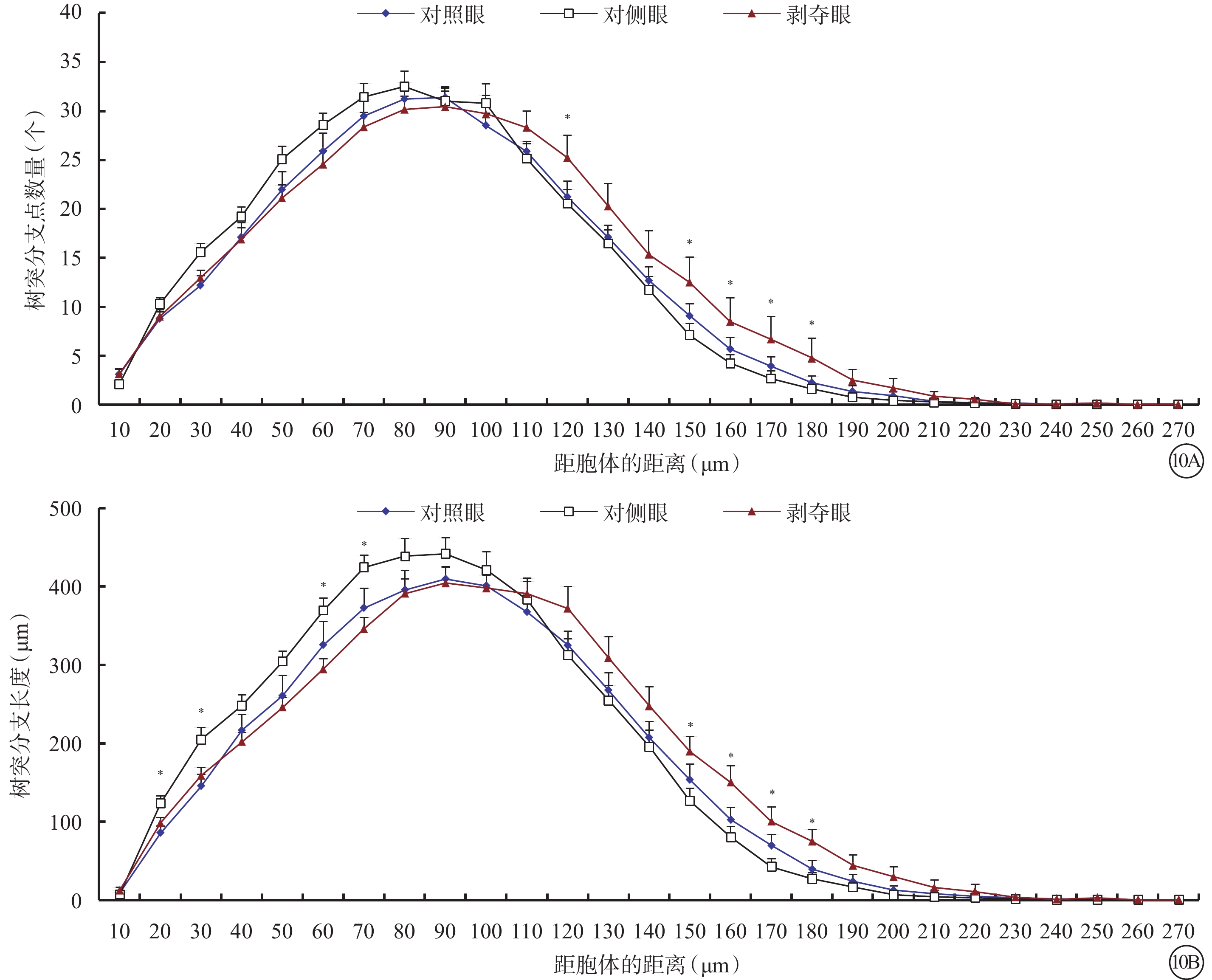 圖10
剝奪眼、對側眼及對照眼Type 2 RGC樹突分布同心圓分析圖。10A示剝奪眼Type 2 RGC距離胞體120~180 μm區域內的樹突分支點數量增多;10B示剝奪眼Type 2 RGC距離胞體150~180 μm區域內樹突分支長度增長,但在20~30 μm、60~70 μm區域內樹突分支長度較對側眼縮短。*P<0.05
圖10
剝奪眼、對側眼及對照眼Type 2 RGC樹突分布同心圓分析圖。10A示剝奪眼Type 2 RGC距離胞體120~180 μm區域內的樹突分支點數量增多;10B示剝奪眼Type 2 RGC距離胞體150~180 μm區域內樹突分支長度增長,但在20~30 μm、60~70 μm區域內樹突分支長度較對側眼縮短。*P<0.05
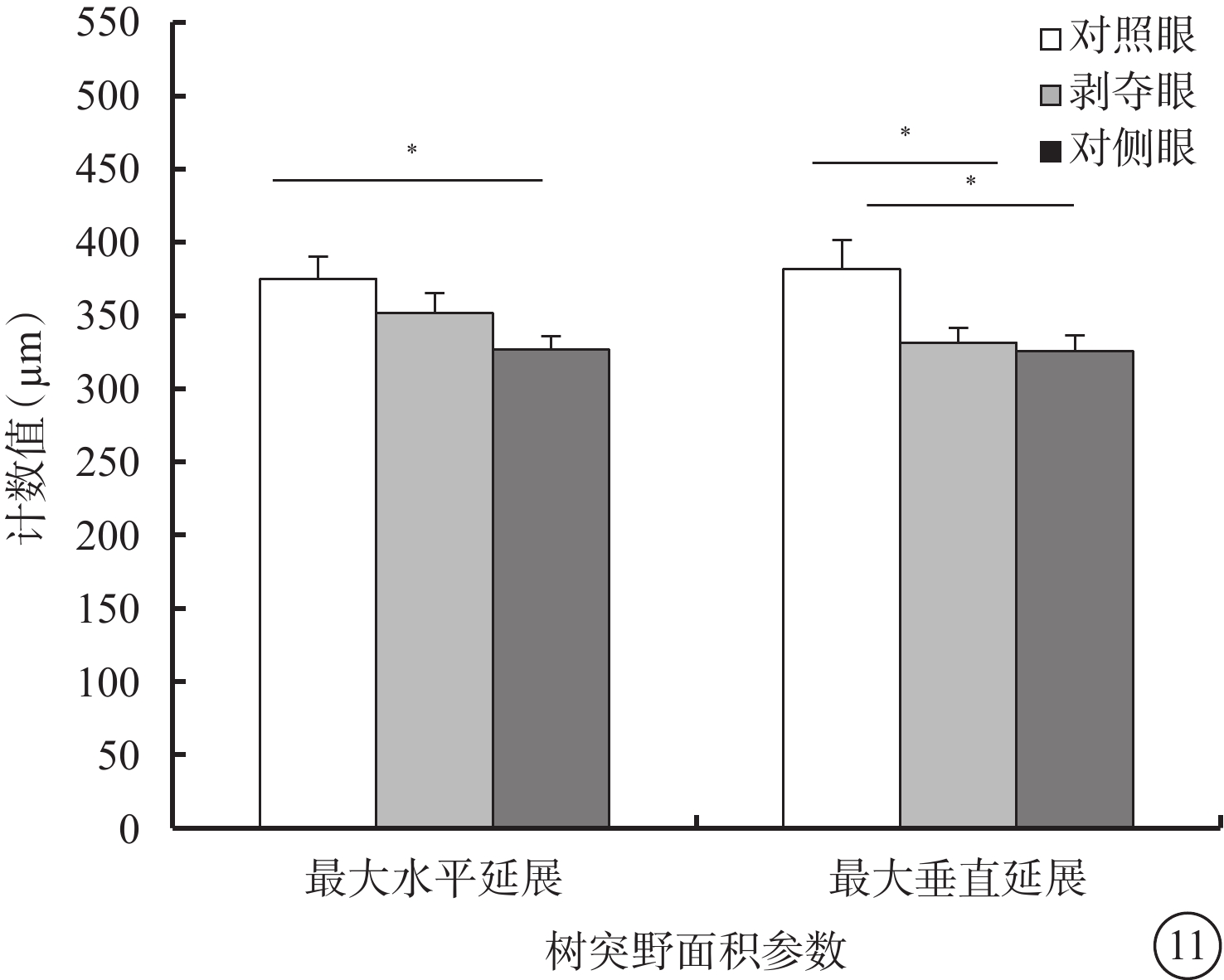
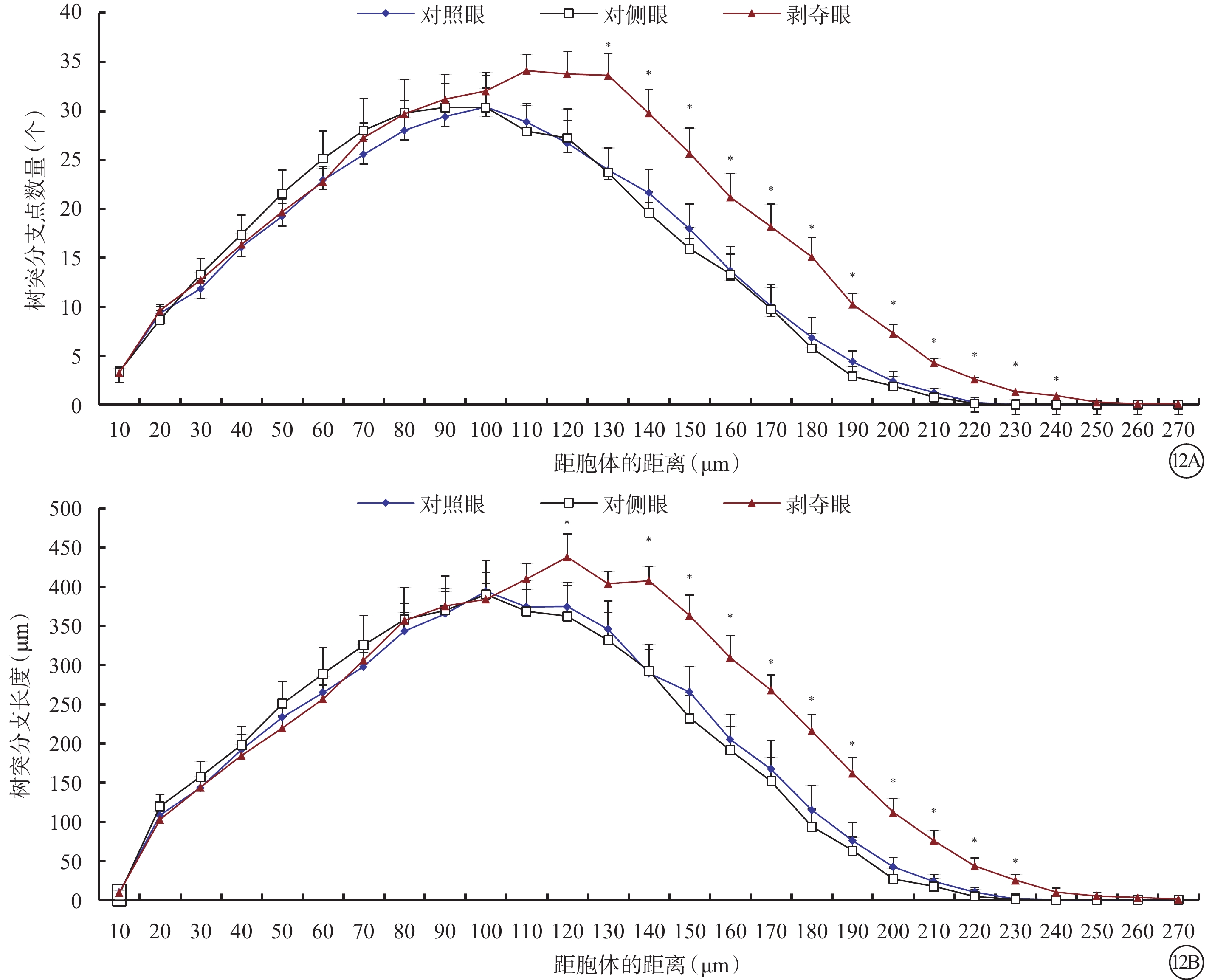
剝奪眼與對側眼Type 3 RGC之間的所有形態參數均無差異(P>0.05)。剝奪眼Type 3 RGC樹突野最大垂直延展較對照眼明顯下降(P=0.035)(圖11)。同心圓分析結果顯示,形覺剝奪明顯增加了130~240 μm區域內Type 3 RGC樹突分支樹突分支點數量,增長了樹突分支長度(圖12)。
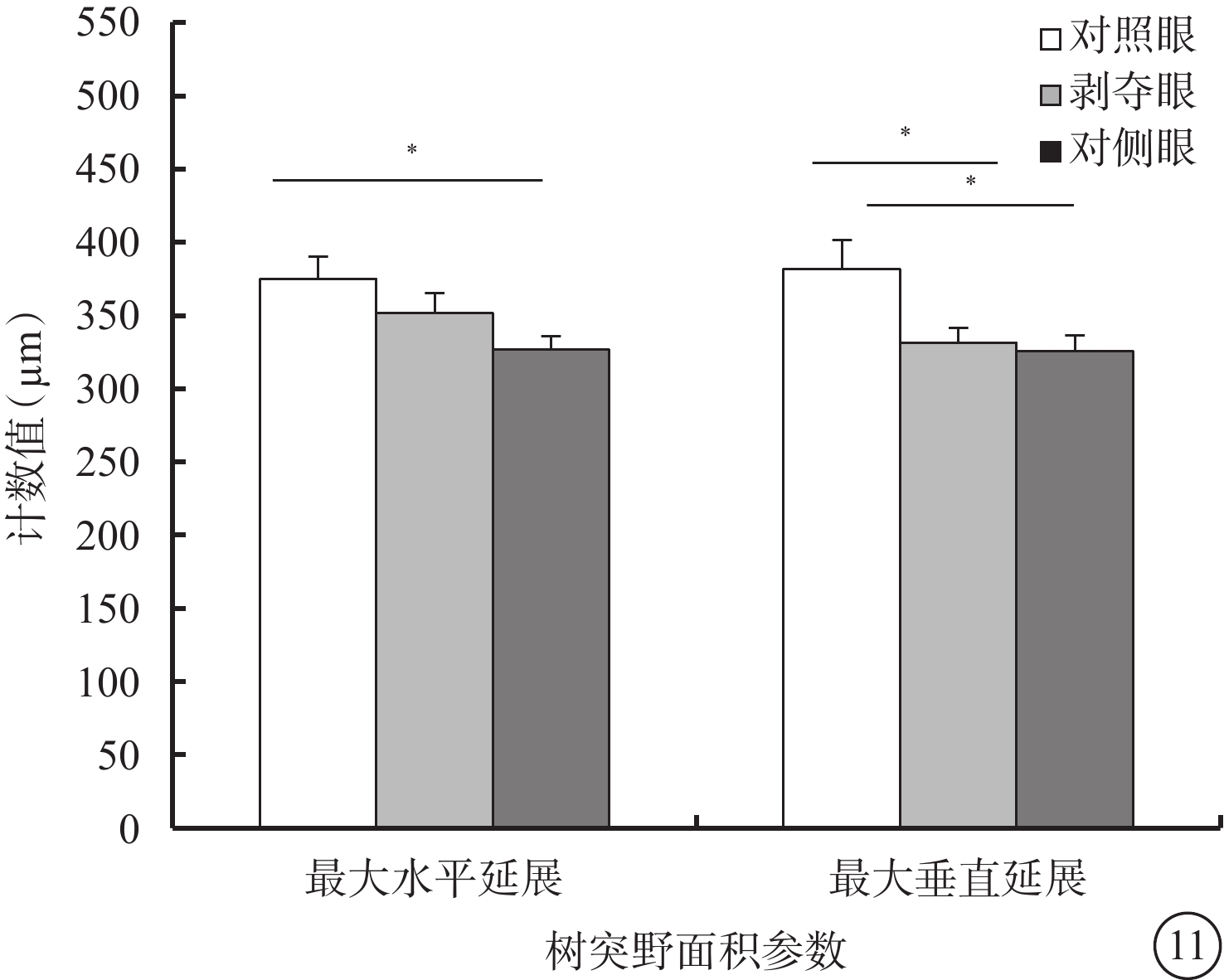 圖11
剝奪眼、對側眼及對照眼Type 3 RGC樹突野面積參數比較。*P<0.05
圖11
剝奪眼、對側眼及對照眼Type 3 RGC樹突野面積參數比較。*P<0.05
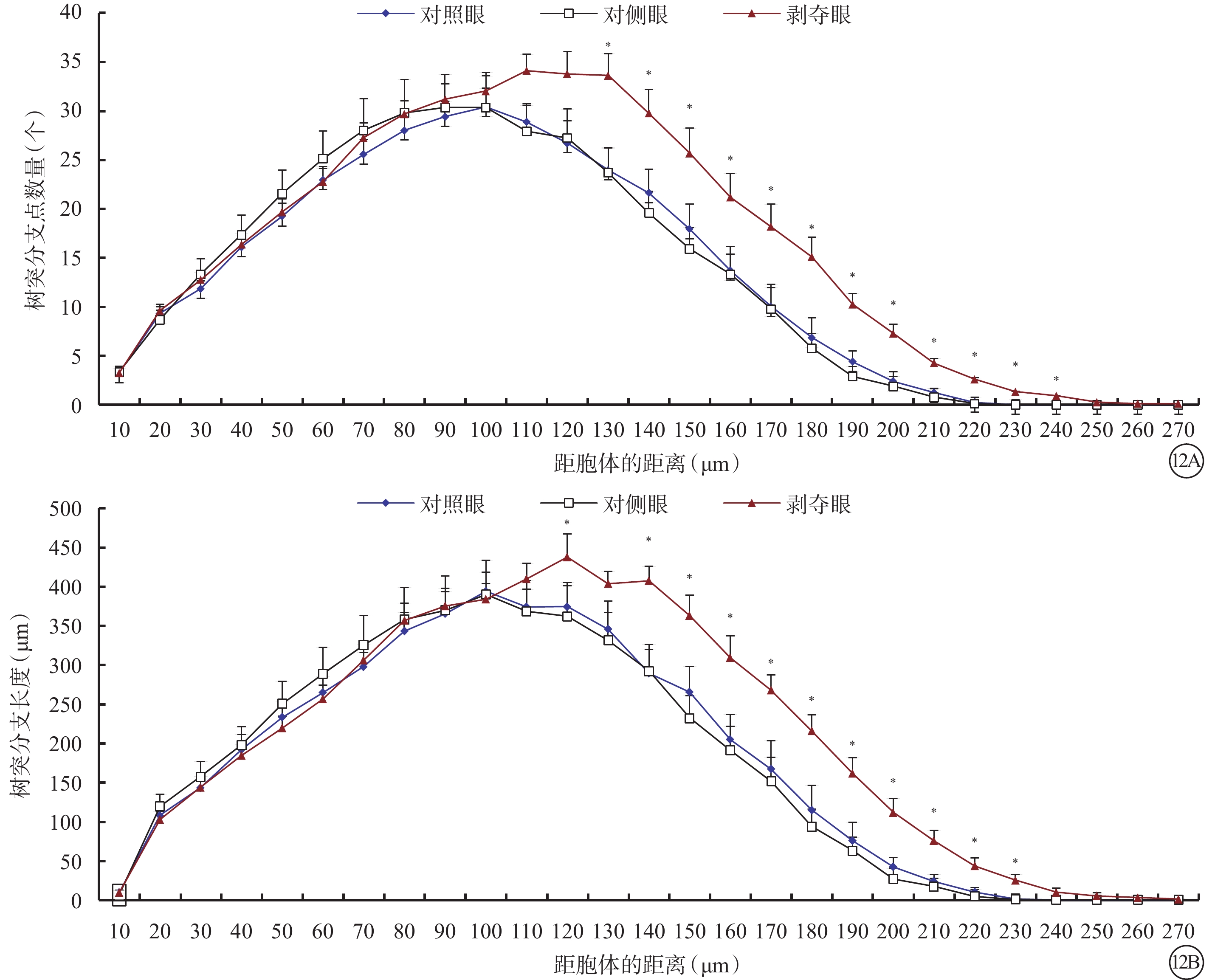 圖12
剝奪眼、對側眼及對照眼Type 3 RGC樹突分布同心圓分析圖。12A示剝奪眼Type 3 RGC距離胞體130~240 μm區域內的樹突分支點數量增多;12B示剝奪眼Type 3 RGC距離胞體120 μm處及150~230 μm區域內樹突分支長度增長。*P<0.05
圖12
剝奪眼、對側眼及對照眼Type 3 RGC樹突分布同心圓分析圖。12A示剝奪眼Type 3 RGC距離胞體130~240 μm區域內的樹突分支點數量增多;12B示剝奪眼Type 3 RGC距離胞體120 μm處及150~230 μm區域內樹突分支長度增長。*P<0.05
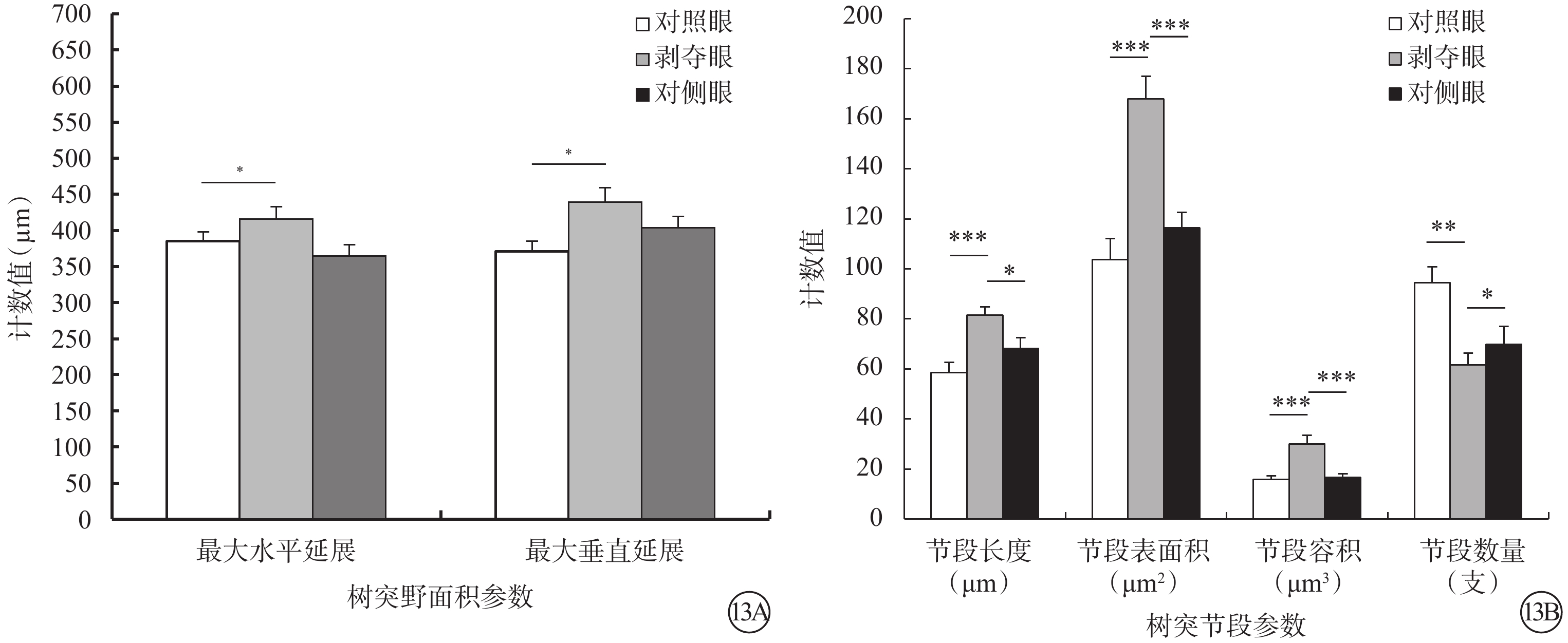
剝奪眼Type 4 RGC樹突野最大水平(F=2.476,P=0.101)、垂直延展(F=4.287,P=0.023)明顯增加,樹突節段長度變長(F=7.482,P=0.002),表面積增大(F=14.814,P=0.000),容積變大(F=11.900,P<0.001),但是樹突節段數量變少(F=7.067,P=0.003)(圖13)。同心圓分析結果顯示,形覺剝奪明顯減少了雙眼120~150 μm區域內以及180 μm處Type 4 RGC樹突分支樹突分支點數量,但對樹突分支長度沒有影響(圖14)。
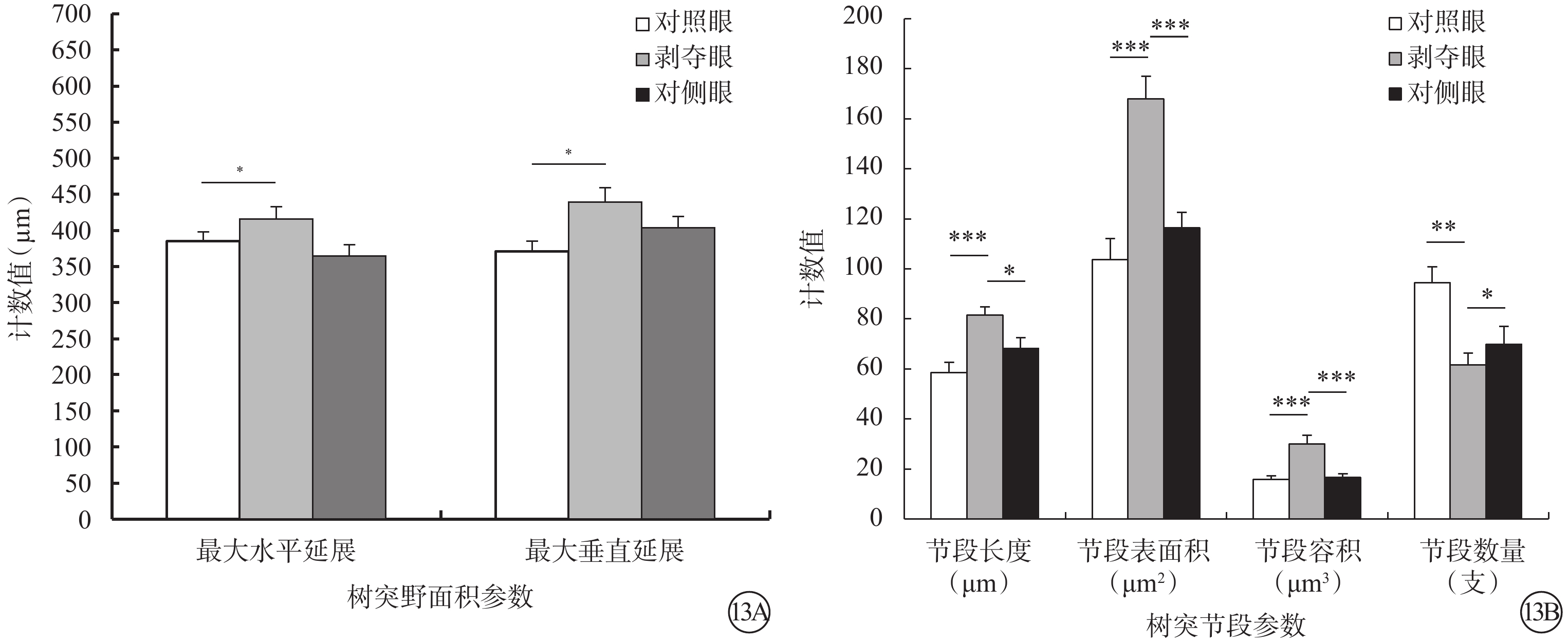 圖13
剝奪眼、對側眼及對照眼Type 4 RGC樹突野面積及樹突節段參數比較。13A示樹突野面積參數;13B示樹突節段參數。*P<0.05,**P<0.01
圖13
剝奪眼、對側眼及對照眼Type 4 RGC樹突野面積及樹突節段參數比較。13A示樹突野面積參數;13B示樹突節段參數。*P<0.05,**P<0.01
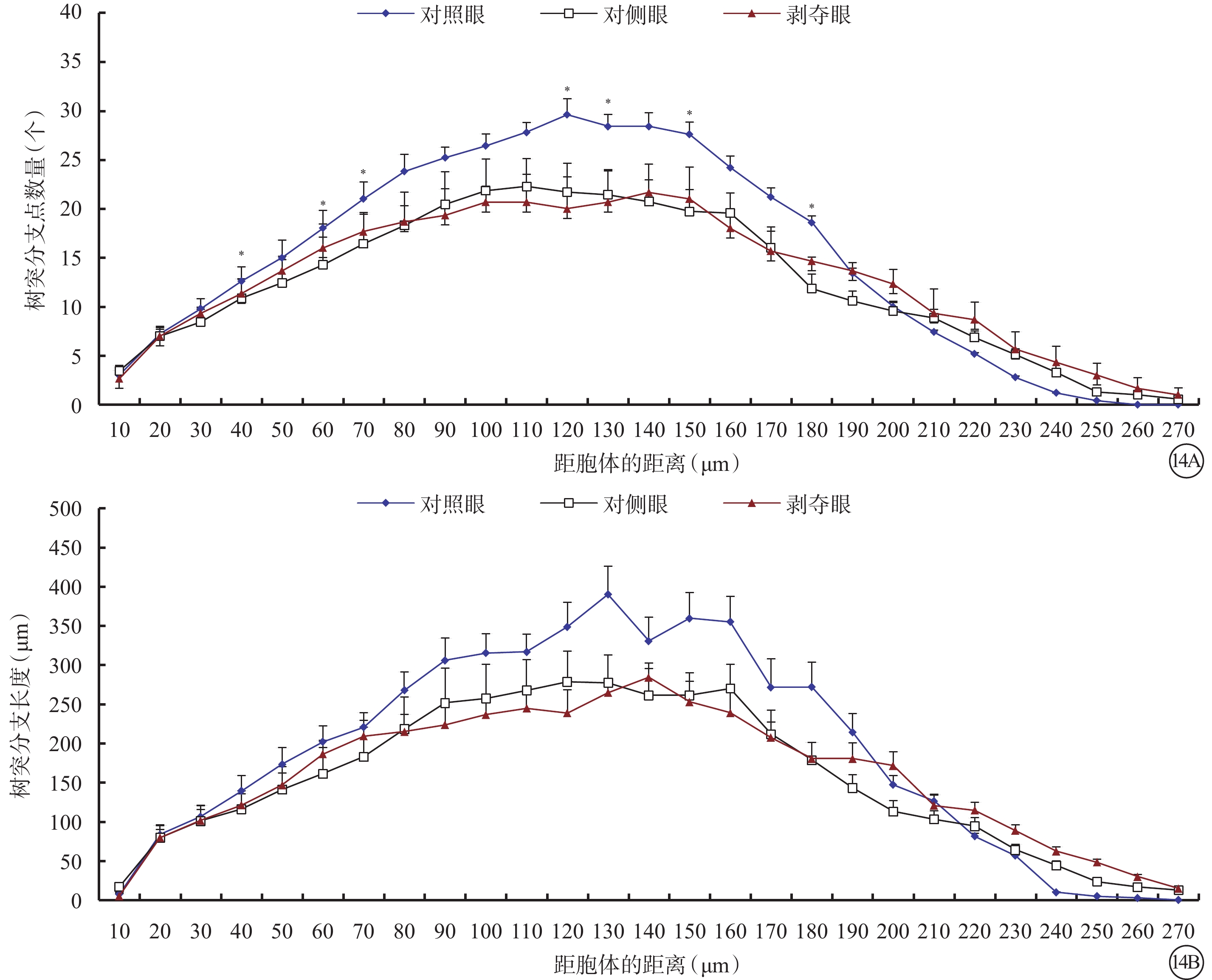 圖14
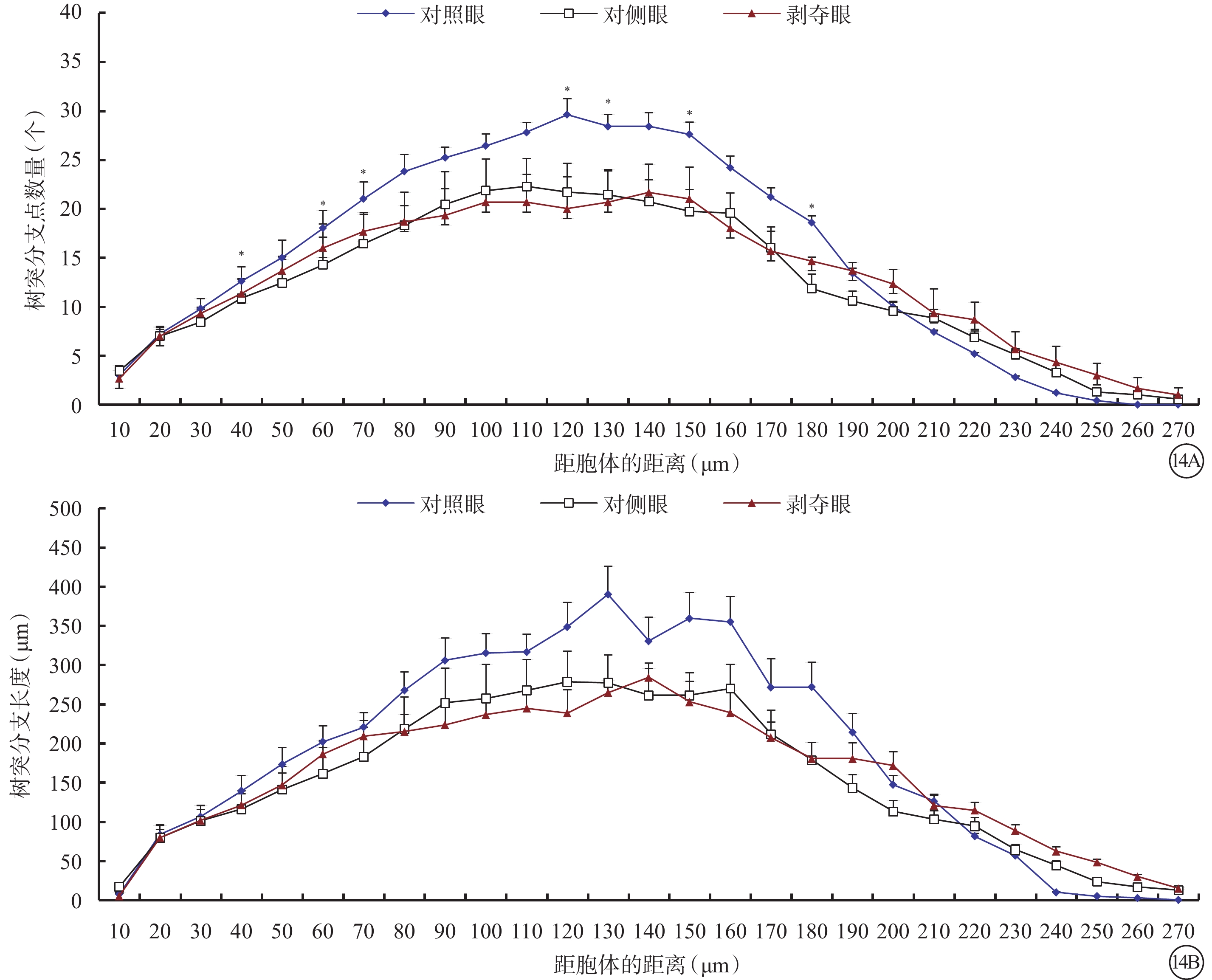
剝奪眼、對側眼及對照眼Type 4 RGC樹突分布同心圓分析圖。14A示形覺剝奪明顯減少了雙眼RGC在120~150 μm區域內以及180 μm處Type 4 RGC樹突分支樹突分支點數量;14B示形覺剝奪對不同同心圓范圍內樹突分支長度沒有影響。*P<0.05
圖14
剝奪眼、對側眼及對照眼Type 4 RGC樹突分布同心圓分析圖。14A示形覺剝奪明顯減少了雙眼RGC在120~150 μm區域內以及180 μm處Type 4 RGC樹突分支樹突分支點數量;14B示形覺剝奪對不同同心圓范圍內樹突分支長度沒有影響。*P<0.05
3 討論
本研究采用轉基因熒光小鼠重構了大量RGC,通過形態學分析軟件并借鑒以往對RGC形態和功能研究文獻將所獲得細胞進行分類。結果顯示,與對側眼與對照眼比較,剝奪眼屈光力向近視方向偏移,RGC數量明顯減少。這說明形覺剝奪對RGC的形態發育有重要影響。除此之外,我們還發現形覺剝奪對4種類型RGC的形態影響各不相同。
Brn3家族是具有POU結構域的轉錄因子,包括Brn3a、Brn3b、Brn3c[9-10]。在視網膜上,Brn3家族只在RGC中有表達,且在完成有絲分裂后,有80%的RGC表達Brn3a。因此我們采用了Brn3a抗體來標記RGC,并進行定量分析。有研究發現,隨著兔子單眼形覺剝奪時間的延長,RGC數量也隨之下降[7]。本研究結果與之一致,在視網膜中心區,剝奪眼RGC數量出現明顯減少的現象。研究發現,阻斷ON型雙極細胞的輸出可以引起ON型RGC減少,提示RGC的生存與其輸入有關[11]。據此我們推測,剝奪眼RGC的數量減少可能與形覺剝奪導致視覺輸入減少有關。此外,視網膜中很多神經遞質的釋放與視覺刺激有關。有學者從出生后23~26 d每日給予小鼠視動刺激,發現其增加了RGC層內腦源性神經生長因子(BDNF)的含量,同時也增加了視網膜多巴胺水平[12]。大部分RGC表達BDNF受體TrkB[13]。已有研究發現,BDNF在缺氧損傷、視神經離斷、高糖時可以對RGC提供神經保護[14-17];在急性眼壓升高時通過TrKB通路對RGC提供抗凋亡作用[18]。和BDNF相似,多巴胺也具有神經保護作用,在發育的視網膜中,多巴胺也具有抗凋亡作用[19]。形覺剝奪在多種物種中可引起視網膜多巴胺水平的下調。多巴胺可以減少大鼠發育視網膜氯胺酮、乙醇誘導的神經元凋亡[20]。因此,我們分析認為,形覺剝奪后RGC數量減少可能與視網膜BDNF和多巴胺水平下降有關。
RGC根據形態、電生理以及基因表達可以分成超過30多種亞型[21-23]。由于本研究僅能從形態入手,缺乏對其電生理以及基因表達的分析,因此難以將RGC明確為某一具體亞型,進而分析其功能。但是通過形態可以進行比較簡單的分類和分析,并與功能較明確的一些亞型進行比較。本研究將200多個重構的RGC分為4種類型。其中Type 1占比最大。它的樹突野較小,樹突支數量少,但是樹突的分支級數最高。這類細胞的樹突節段數量是4類細胞中最多的,但樹突節段短而細。其樹突形態較平直,彎曲度是4類細胞中最小的。形覺剝奪對這類神經元的樹突野大小有輕微增大作用,表現為樹突野的水平延展增大。形覺剝奪后Type 1 RGC樹突彎曲度減少,樹突變得更加平直。這類小樹突野RGC在距離胞體60~100 μm處樹突分布增大。樹突是神經元接受輸入的結構,樹突結構形態和數量的變化提示Type 1 RGC的功能在形覺剝奪的過程發生了改變。Type 1 RGC的形態接近文獻中報道的一些具有小感受野的神經元,如高分辨神經節細胞和局部邊緣探測細胞[24]。兩者對運動視覺刺激的敏感度不同,但均表現出對小視覺刺激物的高敏感度。因此,形覺剝奪過程中,接受小視覺刺激信號的神經通路缺乏刺激,可能在近視的發生發展中發揮了一定的作用。
Type 2、3 RGC樹突野形態上具有一定的相似性。這兩類神經元在形態上與方向選擇性RGC(DSGC)以及α-RGC比較接近。α-RGC的樹突野大小是除ipRGC外最大的,因此Type 3 RGC中可能主要為α-RGC,而Type 2 中DSGC所占的比例可能會更多。形覺剝奪對Type 2、3 RGC的形態影響有所不同,對Type 2的影響主要體現在減少了其在近胞體區域的樹突分布,在20~70 μm區域內樹突分支長度縮短,但在遠胞體段樹突分支數量和長度均有所增加。這提示在形覺剝奪過程中,視覺輸入的減少導致了含有DSGC和α-RGC在內的RGC樹突的重分布。雖然我們的形態學數據不能精確到對真正反映突觸數量的樹突棘的計數,但是樹突分支和長度的重分布依然提示了在形覺剝奪過程中傳遞到這類RGC信號的改變可能是具有異質性的,對近胞體段的輸入改變不同于遠胞體段樹突的輸入改變。形覺剝奪對Type 3 RGC的影響較Type 1、2更為顯著。Type 3 RGC樹突野顯著增加,同時也增加了其遠胞體段樹突分支的數量和長度。以α-RGC為主的Type 3 RGC樹突野的變大會降低視網膜分辨率,樹突分布的改變也提示其輸入在遠胞體段出現了變化。
Type 4 RGC樹突分支數目少,覆蓋范圍大,結構稀疏,與ipRGC形態接近[25-26]。ipRGC主要參與對非形覺的視覺刺激的影響,包括瞳孔反射、晝夜節律等功能。Type 4 RGC樹突水平延展和垂直延展都有所增加。剝奪眼和對側眼的樹突分支點數量以及長度在70~170 μm區域內均明顯減少。變化的趨勢和前三類參與形覺視覺的RGC有所不同,而且是雙眼對稱性的改變。這提示形覺和非形覺RGC的功能在近視的發生發展中可能發揮了不同的作用。
綜上所述,形覺剝奪過程中,視網膜的視覺刺激大幅度缺失,這些缺失的視覺信號包括物體的亮度、顏色、形狀、運動以及非形覺信號等等。視覺刺激的缺失導致RGC減少。本研究發現不同類型的RGC在形覺剝奪后出現的改變很不一致。其中參與形覺信息傳遞的Type 1~3 RGC的變化比較一致,主要表現為神經節距離胞體一定距離的樹突分布變豐富,樹突野范圍表現出變大的趨勢;而參與非形覺信息傳遞的Type 4 RGC則表現出雙眼對稱性改變,樹突分支變稀疏。這提示形覺和非形覺信息在屈光發育中可能具有完全不同的作用,非形覺信息可能具有雙眼效應。
屈光發育關鍵期,合適的視覺輸入是正視化的關鍵因子[1-2]。RGC是視網膜上將視覺信號傳遞到中樞的唯一神經元。不同的視覺信號特征由不同的視網膜神經通路及相應的RGC來承擔[3-5]。視覺信號可分為形覺信號和非形覺信號。形覺信號包括物體的形狀、運動等特征,由傳統的RGC傳遞;非形覺信號參與晝夜節律和瞳孔對光反射等視網膜功能,主要由感光RGC(ipRGC)進行傳遞[3-5]。形覺和非形覺視覺信號在近視的發生發展中均發揮了重要的作用。既往研究發現,超過一半的OFF瞬時性RGC和ON持續性RGC對遠視性和近視性離焦的反應有所不同[6];兔子形覺剝奪過程中RGC的數量減少[7]。這些研究結果證實,RGC功能和數量在近視發生的過程中發揮了作用。RGC的形態是其功能的基礎。比如,局部邊緣探測RGC的樹突野小,分支密集;而ipRGC樹突野大,分支疏松[8]。因此,RGC改變可以反應哪些RGC受形覺剝奪的影響而參與近視過程,以及不同類型的RGC及其所代表的通路是否在近視過程中發揮了不同作用值得深入研究。本研究觀察分析了黃色熒光蛋白(YFP)轉基因小鼠形覺剝奪對不同類型RGC形態的影響,以期為進一步明確近視的視網膜機制提供理論基礎。現將結果報道如下。
1 材料和方法
1.1 實驗動物及其基因型鑒定、實驗設計
本實驗所涉及動物飼養與處理均經溫州醫科大學動物倫理委員會審批通過,并遵循美國視覺與眼科學研究學會關于眼科和視覺研究中對動物的處理原則。60只B6.Cg-Tg(Thy1-YFP)HJrs/J轉基因小鼠(YFP轉基因小鼠)購自美國杰克遜實驗室,繁殖于溫州醫科大學動物實驗室(動物倫理批號WYDW2016-0348)。該小鼠特異性標記10%~30%的RGC。所有小鼠來自YFP轉基因小鼠同窩仔,均在12 h光照/12 h黑暗的照明條件下飼養于標準透光鼠籠中,光照時間為8:00至20:00,環境溫度(22±2)℃。可自由攝食、進水。每3天更換清潔籠具及墊料以保持飼養環境衛生。
剪取小鼠尾巴提取DNA,經PCR后電泳。YFP轉基因小鼠由C57BL/6J為背景,利用轉基因細菌人工染色體技術獲得。在表達胸腺細胞抗原1(Thy1)的基因片段上加上表達YFP的基因片段,雜合子小鼠體內所有表達Thy1的細胞均表達YFP。YFP陽性小鼠Thy1基因片段長度為415 bp,YFP陰性小鼠Thy1基因片段長度為324 bp。
將60只小鼠隨機分為形覺剝奪組及對照組,分別為28、32只。形覺剝奪組小鼠右眼通過遮蓋片進行單眼遮蓋2周作為剝奪眼,同時納入對側眼進行分析。對照組小鼠不作任何處理,取右眼作為對照眼。形覺剝奪前及形覺剝奪后2周,測量兩組小鼠屈光度及眼軸相關參數;采用免疫組織化學染色對視網膜上表達YFP的RGC進行三維重構和形態分析;采用免疫熒光染色對RGC數量進行統計。
1.2 實驗方法
采用自行搭建的紅外偏心攝影驗光儀檢測兩組小鼠的屈光度,每只眼連續測量3次,取平均值。采用Cinescan A/B超聲診斷儀(法國Quantel Medical公司)中的A超探頭進行測量,檢測參數包括前房深度、晶狀體厚度、玻璃體腔深度和眼軸長度。測量前2~3 min用0.5%鹽酸丙美卡因滴眼液進行表面麻醉,每只眼重復測量10次,標準差<0.04 mm,取平均值。
小鼠經5%水合氯醛麻醉后取眼球,分離視網膜,并將其修剪成四葉草形狀。先將視網膜在PBS中清洗3次(10 min/次),然后置于含3% H2O2的PBS中室溫振蕩孵育20 min以去除內源性的過氧化物酶,最后PBS清洗3次(10 min/次)。加入封閉液(5%驢血清+1%牛血清白蛋白+0.2%賴氨酸+1 mol PBS)室溫下封閉2 h后,加入用封閉液稀釋的一抗(抗綠色熒光蛋白,1∶500,美國Novus Biologicals公司)4 ℃孵育2 d后加入用封閉液稀釋的二抗(生物素Biotin標記親和純化驢抗羊IgG二抗,1∶500,美國Jackson Immuno Research公司),室溫孵育2 h,PBS清洗3次(10 min/次)。然后將視網膜置入親和素-生物素復合體(1∶500)4 ℃過夜。PBS清洗后,再經0.1 mol/L三羥甲基氨基甲烷鹽酸鹽(Tris-HCl)清洗5 min。將視網膜置入DAB染色液中避光靜置數分鐘至切片顯色。最后0.1 mol/L Tris-HCl清洗10 min,PBS清洗2次(10 min/次),封片。
封片后的載玻片靜置晾干。在明場10倍鏡下,挑選顯色清晰、結構完整的RGC,先描繪視網膜的外輪廓。在63倍水鏡下,使用Neurolucida分析系統在線神經元三維重構系統重構目標神經元的三維立體結構,包括胞體、樹突和軸突(圖1A)。利用Neurolucida Explorer分析軟件對重構的RGC三維結構進行定量分析對比,包括胞體的周長及面積、樹突數量、樹突長度、樹突表面積、樹突體積、樹突的水平及垂直延展距離、樹突分級數、彎曲度及分支角度;同心圓分析獲得樹突分支點數目及分支長度等參數在內的23個形態學參數(圖1B~1G)。
圖1
部分形態學參數分析示意圖
小鼠頸椎脫臼后取眼球,4%多聚甲醛溶液固定20 min后取視網膜并修剪成四葉草形狀。視網膜經0.1 mol PBS中清洗后加入封閉液(6%驢血清+1%牛血清白蛋白+ 0.2% Triton X-100,pH=7.4)室溫封閉2 h。然后加入一抗(Brn3a抗體,sc-31984,1:50,美國Santa Cruz公司),4 ℃孵育2 d,加入二抗(生物素Biotin標記親和純化驢抗羊IgG二抗,1:400,美國Jackson immuno Research公司)孵育2 h。PBS清洗3次(10 min/次),封片。利用共聚焦顯微鏡(Imager-Z1,德國Zeiss公司)在視網膜四葉草的近端和遠端兩個視野拍照,共取8個視野進行分析。計數DAPI標記的所有RCG層細胞和Brn3a抗體標記的RGC。
1.3 統計學方法
采用SPSS19.0軟件進行統計分析,數據用均數±標準差()表示。形覺剝奪組剝奪眼及其對側眼、對照眼之間各參數比較采用單因素方差分析,并用最小顯著差法校正。組內左右眼之間各參數比較采用配對 t 檢驗。不同類型RGC占所有重構RGC的比例比較采用 χ2 檢驗。P<0.05為差異有統計學意義。
2 結果
2.1 形覺剝奪對屈光度、眼軸相關參數及RGC數量的影響
形覺剝奪前,形覺剝奪組剝奪眼、對側眼以及對照眼屈光度(F=0.849,P=0.436)和前房深度(F=0.197,P=0.822)、玻璃體腔深度(F=0.470,P=0.629)、眼軸長度(F=0.024,P=0.102)等眼軸相關參數比較,差異無統計學意義。形覺剝奪后2周,形覺剝奪組剝奪眼較對照眼出現了顯著的近視改變(F=15.009,P<0.001),同時伴有玻璃體腔深度加深(F=3.360,P=0.047)、眼軸延長(F=5.011,P=0.013);但前房深度和晶狀體厚度無明顯變化(P>0.05)(表1,圖2)。
)
圖2
形覺剝奪后2周形覺剝奪組剝奪眼、對側眼及對照眼屈光度、眼軸相關參數比較。2A示屈光度;2B示玻璃體腔深度;2C示眼軸長度。*P<0.05,**P<0.001
形覺剝奪組剝奪眼(n=4)、對側眼(n=3)以及對照眼(n=4)視網膜后極部RGC數量分別為(324±23)、(279±15)、(377±23)個,視網膜周邊部RGC數量分別為(270±31)、(288±8)、(310±47)個。在視網膜后極部,形覺剝奪組剝奪眼RGC數量較對側眼及對照組明顯減少,差異有統計學意義(F=4.769,P=0.035);但在周邊區,差異無統計學意義(F=1.517,P=0.266)。
2.2 RGC的類型及形態
根據樹突野形態,三維重構的RGC可分為Type 1、Type 2、Type 3、Type 4共4種類型(圖3)。4種類型的RGC占比無明顯差異(χ2=6.853,P=0.335,各類型調整殘差≤2)(圖4)。
圖3
不同類型RGC形態示意圖。3A示Type 1;3B示Type 2;3C示Type 3;3D示Type 4
圖4
不同類型RGC在視網膜上的分布及其占各組重構細胞的比例。4A示4種類型RGC在視網膜上的分布示意圖,藍色代表Type 1,白色代表Type 2,紅色代表Type 3,綠色代表Type 4;4B示4種類型RGC占各組重構細胞的比例
Type 1 RGC具有小而緊湊的樹突野;樹突支較少且短;樹突支分支級數、樹突節段多但短。與Type 1 RGC比較,Type 2 RGC樹突野更大;樹突分布較稀疏;樹突支數量更多且更長;樹突支分支級數、樹突節段數較少,但長度更長。與Type 2 RGC比較,Type 3 RGC樹突野更大,形態相似;樹突分布更豐富;樹突支數量更多且更長;樹突支分支級數、樹突節段數較少,但長度更長。Type 4 RGC樹突野形態是4種類型RGC中最大的;樹突分布最為稀疏;樹突支數量與Type 2、3 RGC相仿,長度略長;樹突支分支級數與Type 3 RGC接近;樹突節段數量與Type 2、3 RGC接近,但長度更長(表2,圖5)。
)
圖5
4種類型RGC樹突野、樹突支、樹突分支以及樹突節段等參數比較。5A示樹突野參數;5B示樹突支參數;5C示樹突分支參數;5D示樹突節段參數。*P<0.05
同心圓分析結果顯示,Type 1、Type 2、Type 3、Type 4 RGC樹突分支最多的區域分別集中在距離胞體50、90、100、130 μm處,分別在距離胞體140、200、210、240 μm處基本不再發出新的分支(圖6)。Type 3 RGC樹突分支最多的區域與Type 2 RGC接近,但在距離胞體110 μm以內樹突分支數量和長度較Type 2 RGC明顯少和短,在110 μm以外則樹突分支數量和長度均較Type 2 RGC更多。
圖6
4種類型RGC同心圓分析圖。6A示離胞體不同距離內樹突分支點的數量;6B示離胞體不同距離內樹突分支長度。*P<0.05
2.3 形覺剝奪對4種類型RGC形態的影響
形覺剝奪對Type 1 RGC樹突野大小影響不大,但明顯增加了樹突野最大水平延展,降低了樹突彎曲度。與對側眼及對照眼比較,剝奪眼Type 1 RGC樹突野最大水平延展明顯增加(F=4.312,P=0.019),樹突彎曲度明顯下降(F=6.608,P=0.003),差異均有統計學意義(圖7)。同心圓分析結果顯示,形覺剝奪明顯增加了Type 1 RGC距離胞體50~90 μm區域內的樹突分支點數量,增長了樹突分支長度(圖8)。
圖7
剝奪眼、對側眼及對照眼Type 1 RGC樹突野面積及樹突彎曲度參數比較。7A示樹突野面積參數;7B示樹突彎曲度參數。*P<0.05,**P<0.01,***P<0.001
圖8
剝奪眼、對側眼及對照眼Type 1 RGC樹突分布同心圓分析圖。8A示剝奪眼Type 1 RGC距離胞體50~90 μm區域內的樹突分支點數量增多;8B示剝奪眼Type 1 RGC距離胞體50~90 μm區域內樹突分支長度增長。*P<0.05
和Type1 RGC接近,形覺剝奪對Type 2 RGC的樹突野面積(F=1.780,P=0.180)以及最大水平延展(F=2.722,P=0.077)沒有明顯影響,但增加了其最大垂直延展(F=4.704,P=0.014)。同時,形覺剝奪減小了樹突分支角(F=5.180,P=0.009)以及樹突樣條曲線角度數(F=5.672,P=0.006)。剝奪眼Type 2 RGC樹突野最大垂直延展較對側眼明顯增加(P=0.012),但與對照眼無明顯差異(P=0.728)。剝奪眼Type 2 RGC樹突彎曲度和對側眼及對照眼相比均無明顯改變(F=2.606,P=0.085);但樹突分支角(P=0.046)、樹突樣條曲線角(P=0.026)較對側眼明顯變小(圖9)。同心圓分析結果顯示,形覺剝奪明顯縮短了20~30 μm及60~70 μm區域內Type 2 RGC樹突分支長度;增加了120~180 μm區域內Type 2 RGC樹突分支點數量以及150~180 μm區域內樹突分支長度(圖10)。
圖9
剝奪眼、對側眼及對照眼Type 2 RGC樹突野面積及樹突彎曲度參數比較。9A示樹突野面積參數;9B示樹突彎曲度參數。*P<0.05,**P<0.01
圖10
剝奪眼、對側眼及對照眼Type 2 RGC樹突分布同心圓分析圖。10A示剝奪眼Type 2 RGC距離胞體120~180 μm區域內的樹突分支點數量增多;10B示剝奪眼Type 2 RGC距離胞體150~180 μm區域內樹突分支長度增長,但在20~30 μm、60~70 μm區域內樹突分支長度較對側眼縮短。*P<0.05
剝奪眼與對側眼Type 3 RGC之間的所有形態參數均無差異(P>0.05)。剝奪眼Type 3 RGC樹突野最大垂直延展較對照眼明顯下降(P=0.035)(圖11)。同心圓分析結果顯示,形覺剝奪明顯增加了130~240 μm區域內Type 3 RGC樹突分支樹突分支點數量,增長了樹突分支長度(圖12)。
圖11
剝奪眼、對側眼及對照眼Type 3 RGC樹突野面積參數比較。*P<0.05
圖12
剝奪眼、對側眼及對照眼Type 3 RGC樹突分布同心圓分析圖。12A示剝奪眼Type 3 RGC距離胞體130~240 μm區域內的樹突分支點數量增多;12B示剝奪眼Type 3 RGC距離胞體120 μm處及150~230 μm區域內樹突分支長度增長。*P<0.05
剝奪眼Type 4 RGC樹突野最大水平(F=2.476,P=0.101)、垂直延展(F=4.287,P=0.023)明顯增加,樹突節段長度變長(F=7.482,P=0.002),表面積增大(F=14.814,P=0.000),容積變大(F=11.900,P<0.001),但是樹突節段數量變少(F=7.067,P=0.003)(圖13)。同心圓分析結果顯示,形覺剝奪明顯減少了雙眼120~150 μm區域內以及180 μm處Type 4 RGC樹突分支樹突分支點數量,但對樹突分支長度沒有影響(圖14)。
圖13
剝奪眼、對側眼及對照眼Type 4 RGC樹突野面積及樹突節段參數比較。13A示樹突野面積參數;13B示樹突節段參數。*P<0.05,**P<0.01
圖14
剝奪眼、對側眼及對照眼Type 4 RGC樹突分布同心圓分析圖。14A示形覺剝奪明顯減少了雙眼RGC在120~150 μm區域內以及180 μm處Type 4 RGC樹突分支樹突分支點數量;14B示形覺剝奪對不同同心圓范圍內樹突分支長度沒有影響。*P<0.05
3 討論
本研究采用轉基因熒光小鼠重構了大量RGC,通過形態學分析軟件并借鑒以往對RGC形態和功能研究文獻將所獲得細胞進行分類。結果顯示,與對側眼與對照眼比較,剝奪眼屈光力向近視方向偏移,RGC數量明顯減少。這說明形覺剝奪對RGC的形態發育有重要影響。除此之外,我們還發現形覺剝奪對4種類型RGC的形態影響各不相同。
Brn3家族是具有POU結構域的轉錄因子,包括Brn3a、Brn3b、Brn3c[9-10]。在視網膜上,Brn3家族只在RGC中有表達,且在完成有絲分裂后,有80%的RGC表達Brn3a。因此我們采用了Brn3a抗體來標記RGC,并進行定量分析。有研究發現,隨著兔子單眼形覺剝奪時間的延長,RGC數量也隨之下降[7]。本研究結果與之一致,在視網膜中心區,剝奪眼RGC數量出現明顯減少的現象。研究發現,阻斷ON型雙極細胞的輸出可以引起ON型RGC減少,提示RGC的生存與其輸入有關[11]。據此我們推測,剝奪眼RGC的數量減少可能與形覺剝奪導致視覺輸入減少有關。此外,視網膜中很多神經遞質的釋放與視覺刺激有關。有學者從出生后23~26 d每日給予小鼠視動刺激,發現其增加了RGC層內腦源性神經生長因子(BDNF)的含量,同時也增加了視網膜多巴胺水平[12]。大部分RGC表達BDNF受體TrkB[13]。已有研究發現,BDNF在缺氧損傷、視神經離斷、高糖時可以對RGC提供神經保護[14-17];在急性眼壓升高時通過TrKB通路對RGC提供抗凋亡作用[18]。和BDNF相似,多巴胺也具有神經保護作用,在發育的視網膜中,多巴胺也具有抗凋亡作用[19]。形覺剝奪在多種物種中可引起視網膜多巴胺水平的下調。多巴胺可以減少大鼠發育視網膜氯胺酮、乙醇誘導的神經元凋亡[20]。因此,我們分析認為,形覺剝奪后RGC數量減少可能與視網膜BDNF和多巴胺水平下降有關。
RGC根據形態、電生理以及基因表達可以分成超過30多種亞型[21-23]。由于本研究僅能從形態入手,缺乏對其電生理以及基因表達的分析,因此難以將RGC明確為某一具體亞型,進而分析其功能。但是通過形態可以進行比較簡單的分類和分析,并與功能較明確的一些亞型進行比較。本研究將200多個重構的RGC分為4種類型。其中Type 1占比最大。它的樹突野較小,樹突支數量少,但是樹突的分支級數最高。這類細胞的樹突節段數量是4類細胞中最多的,但樹突節段短而細。其樹突形態較平直,彎曲度是4類細胞中最小的。形覺剝奪對這類神經元的樹突野大小有輕微增大作用,表現為樹突野的水平延展增大。形覺剝奪后Type 1 RGC樹突彎曲度減少,樹突變得更加平直。這類小樹突野RGC在距離胞體60~100 μm處樹突分布增大。樹突是神經元接受輸入的結構,樹突結構形態和數量的變化提示Type 1 RGC的功能在形覺剝奪的過程發生了改變。Type 1 RGC的形態接近文獻中報道的一些具有小感受野的神經元,如高分辨神經節細胞和局部邊緣探測細胞[24]。兩者對運動視覺刺激的敏感度不同,但均表現出對小視覺刺激物的高敏感度。因此,形覺剝奪過程中,接受小視覺刺激信號的神經通路缺乏刺激,可能在近視的發生發展中發揮了一定的作用。
Type 2、3 RGC樹突野形態上具有一定的相似性。這兩類神經元在形態上與方向選擇性RGC(DSGC)以及α-RGC比較接近。α-RGC的樹突野大小是除ipRGC外最大的,因此Type 3 RGC中可能主要為α-RGC,而Type 2 中DSGC所占的比例可能會更多。形覺剝奪對Type 2、3 RGC的形態影響有所不同,對Type 2的影響主要體現在減少了其在近胞體區域的樹突分布,在20~70 μm區域內樹突分支長度縮短,但在遠胞體段樹突分支數量和長度均有所增加。這提示在形覺剝奪過程中,視覺輸入的減少導致了含有DSGC和α-RGC在內的RGC樹突的重分布。雖然我們的形態學數據不能精確到對真正反映突觸數量的樹突棘的計數,但是樹突分支和長度的重分布依然提示了在形覺剝奪過程中傳遞到這類RGC信號的改變可能是具有異質性的,對近胞體段的輸入改變不同于遠胞體段樹突的輸入改變。形覺剝奪對Type 3 RGC的影響較Type 1、2更為顯著。Type 3 RGC樹突野顯著增加,同時也增加了其遠胞體段樹突分支的數量和長度。以α-RGC為主的Type 3 RGC樹突野的變大會降低視網膜分辨率,樹突分布的改變也提示其輸入在遠胞體段出現了變化。
Type 4 RGC樹突分支數目少,覆蓋范圍大,結構稀疏,與ipRGC形態接近[25-26]。ipRGC主要參與對非形覺的視覺刺激的影響,包括瞳孔反射、晝夜節律等功能。Type 4 RGC樹突水平延展和垂直延展都有所增加。剝奪眼和對側眼的樹突分支點數量以及長度在70~170 μm區域內均明顯減少。變化的趨勢和前三類參與形覺視覺的RGC有所不同,而且是雙眼對稱性的改變。這提示形覺和非形覺RGC的功能在近視的發生發展中可能發揮了不同的作用。
綜上所述,形覺剝奪過程中,視網膜的視覺刺激大幅度缺失,這些缺失的視覺信號包括物體的亮度、顏色、形狀、運動以及非形覺信號等等。視覺刺激的缺失導致RGC減少。本研究發現不同類型的RGC在形覺剝奪后出現的改變很不一致。其中參與形覺信息傳遞的Type 1~3 RGC的變化比較一致,主要表現為神經節距離胞體一定距離的樹突分布變豐富,樹突野范圍表現出變大的趨勢;而參與非形覺信息傳遞的Type 4 RGC則表現出雙眼對稱性改變,樹突分支變稀疏。這提示形覺和非形覺信息在屈光發育中可能具有完全不同的作用,非形覺信息可能具有雙眼效應。