引用本文: 張哲, 劉巨平, 東莉潔, 劉竹青, 黃亮瑜, 蘇睿虹, 趙今稚, 張曉敏, 李筱榮. 高糖狀態下視網膜血管內皮細胞基因表達譜的RNA-Seq分析. 中華眼底病雜志, 2018, 34(4): 377-381. doi: 10.3760/cma.j.issn.1005-1015.2018.04.014 復制
研究發現,糖尿病視網膜病變(DR)初期就已出現血管內皮細胞功能異常,且隨著血糖升高,其受損進一步加重[1]。血糖濃度增高可抑制血管舒張,在高糖狀態下,視網膜血管的舒張功能減退,血流速度下降[2]。但鑒于高糖對血管內皮細胞的干預是一個多方面、多層次的復雜過程,我們有必要利用更為先進的檢測方法,在更廣泛的檢測范圍內了解高糖作用下血管內皮細胞改變的完整信息,以期對疾病的后續預防與治療提供有力的依據。RNA轉錄組分析是利用新一代高通量測序技術對全轉錄組測序研究的技術,轉錄組測序的研究對象為特定細胞在某一功能狀態下所能轉錄出來的所有RNA的總和[3]。轉錄組是連接基因組遺傳信息與生物功能蛋白質組的必然紐帶,也是基因功能及結構研究的基礎和出發點。通過新一代高通量測序,能夠全面快速地獲得某一物種特定組織或器官在特定狀態下的幾乎所有轉錄本序列信息,已廣泛應用于基礎研究、臨床診斷和藥物研發等領域[4, 5]。本研究基于RNA轉錄組分析技術,研究高糖狀態對視網膜血管內皮細胞的影響,旨在全面完整的挖掘并分析高糖狀態對血管內皮細胞影響的生物學信息,從而為DR的治療提供更為準確的切入點。
1 材料和方法
1.1 主要材料、細胞培養及實驗分組
恒河猴視網膜血管內皮細胞(RF-6A),由天津醫科大學眼科醫院研究所提供。磷酸鹽緩沖液(PBS)、RPMI-1640培養液(上海源培生物科技股份有限公司),胰蛋白酶、胎牛血清、青鏈霉素、谷氨酰胺(美國Gibco公司)。6孔板、96孔板(美國Life Technologies公司),Trizol試劑盒(美國Invitrogen公司)。葡萄糖(美國Sigma公司)。
RF-6A細胞采用含4.5 ml胎牛血清、0.45 ml雙抗、0.45 ml谷氨酰胺的RPMI-1640培養基于37℃、5%的CO2培養箱中培養,3~5 d傳代1次。取對數生長期細胞用于實驗。實驗分為對照組、高糖組進行,兩組細胞分別采用5、25 mmol/L的葡萄糖持續培養48 h。
1.2 轉錄組文庫測序
轉錄組文庫測序的詳細方法參照文獻[6]。提取不同分組細胞中的總RNA,并對所提取的總RNA的質量進行多方面的質控,以確保后續處理的數據質量合格。隨后在確定總RNA樣品檢測合格后,應用磁珠對poly A尾的mRNA進行富集及片段化,并依據NEB普通建庫方式及鏈特異性建庫方式進行建庫。文庫構建完成后,在經熒光計進行初步定量的基礎之上,應用熒光定量逆轉錄來準確定量文庫有效濃度(文庫有效濃度高于2 nmol/L),以保證文庫質量。庫檢合格后,將不同文庫按照有效濃度及目標下機數據量的需求進行數據匯總分析,經Illumina HiSeq/MiSeq測序儀通過捕獲熒光信號并應用計算機軟件將光信號轉化為測序峰,從而獲得待測片段的序列信息。
1.3 差異表達基因及功能富集分析
將得到的表達譜進行差異表達分析。采用軟件Bioconductor中的edgeR函數做差異表達分析,edgeR函數假設測序reads的計數對于每個基因來說是負二項分布,基于此理論分布來做假設檢驗。并將差異顯著的基因(P≤0.001)提取相應的蛋白質序列進行GO功能、Pathway顯著性富集分析。
應用Blast軟件比對基因序列到NR庫和KEGG數據庫,隨后提取所有基因的GO、Pathway注釋信息。根據差異表達基因列表,應用軟件Bioconductor中的topGO函數和超幾何檢驗對差異表達基因進行Pathway、GO功能顯著性富集分析。對得到的差異基因大數據進行篩查,篩除Log-2到Log2區間內無意義的數據,將該數據按P值大小逐一從數據庫中找出對應基因,查閱相關文獻,了解該基因在不同組織中發揮的作用及作用機制,根據基因的作用途徑,完善顯著差異基因間的關系。
2 結果
所有樣品在illumina測序平臺的轉錄組測序后,得到對照組和高糖組總的reads數分別有50 757 454和49 120 408條;過濾質量較低的數據,得到clean reads數所占比例分別為94.30%、95.14%(表1)。
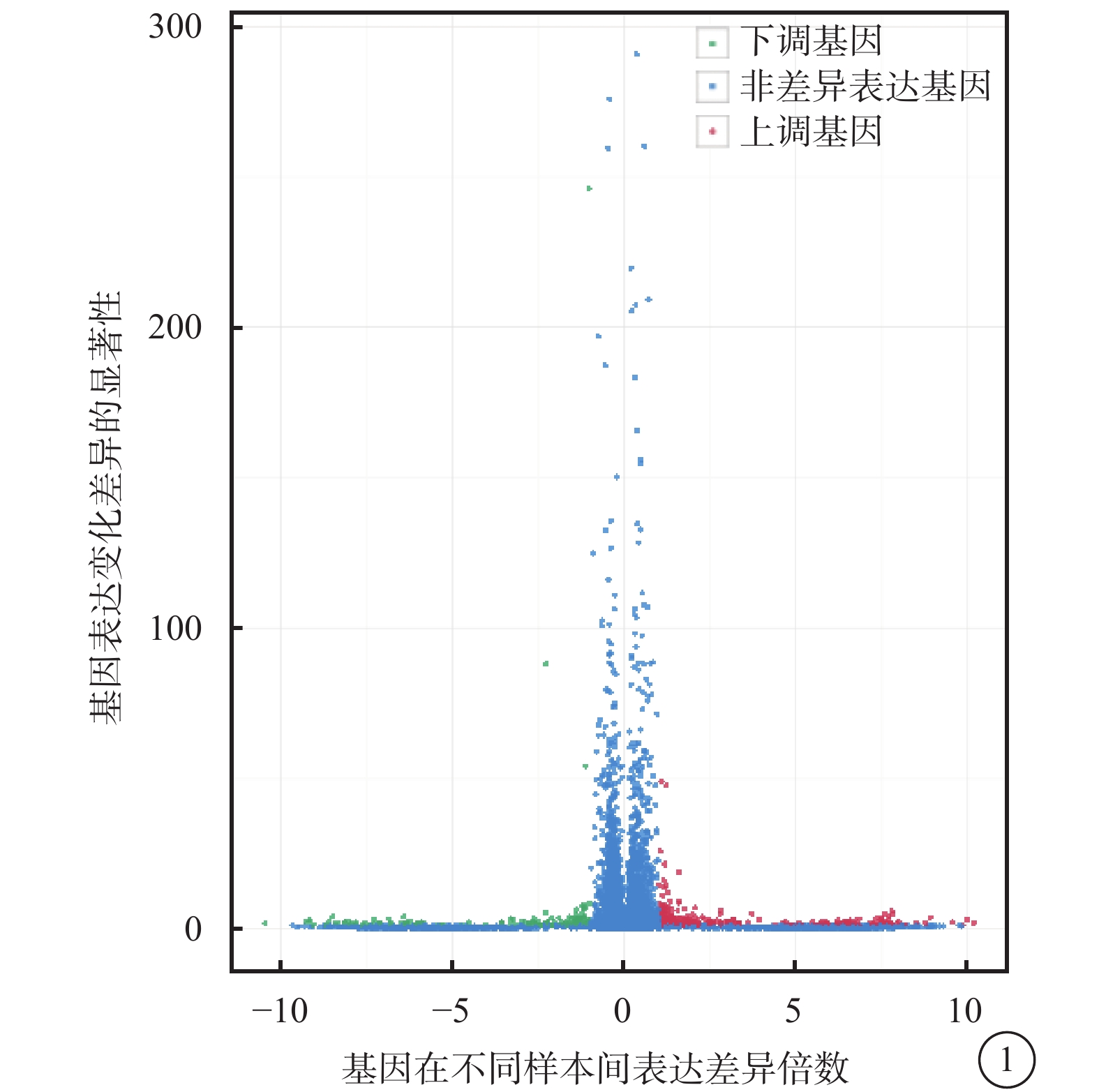
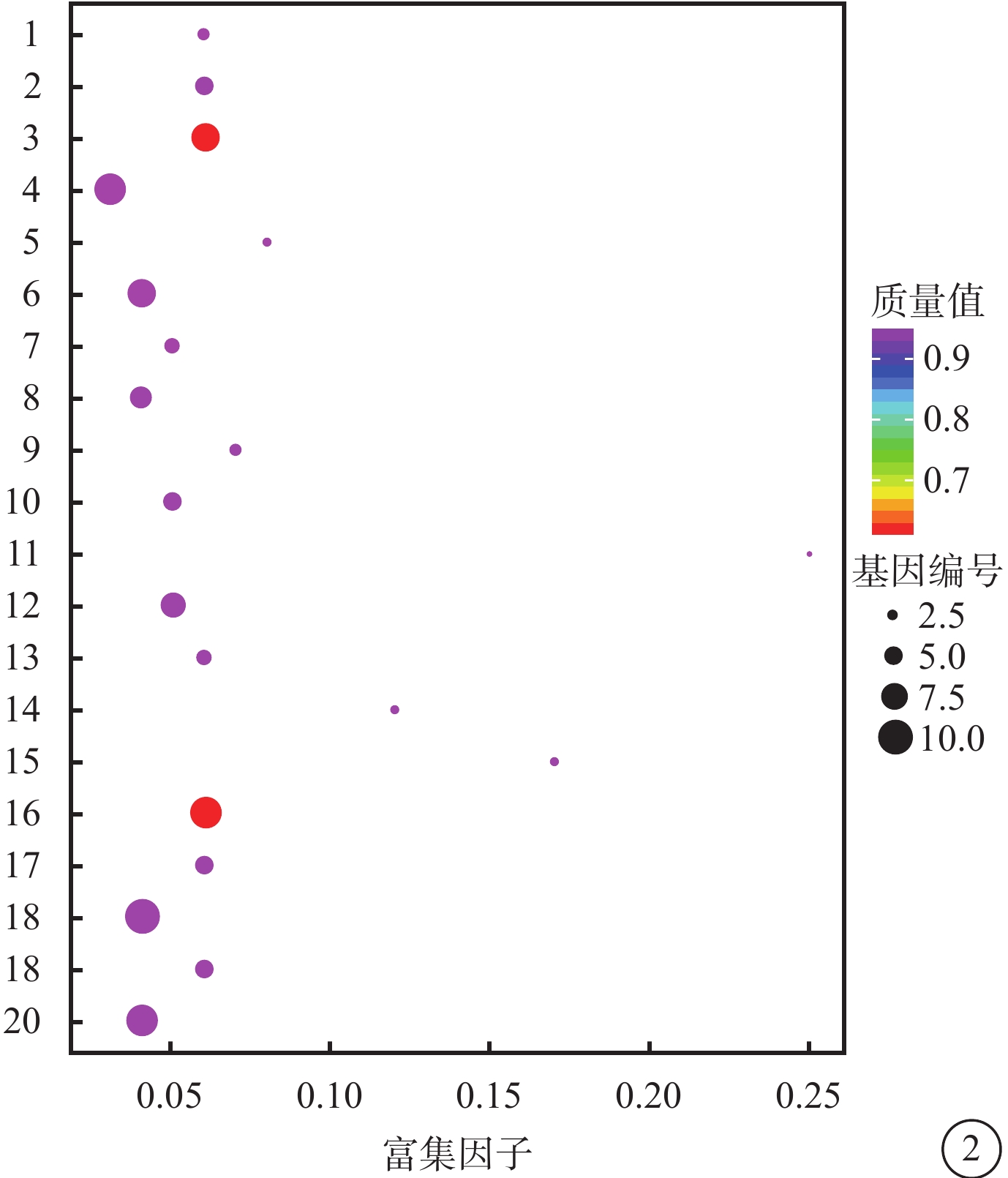
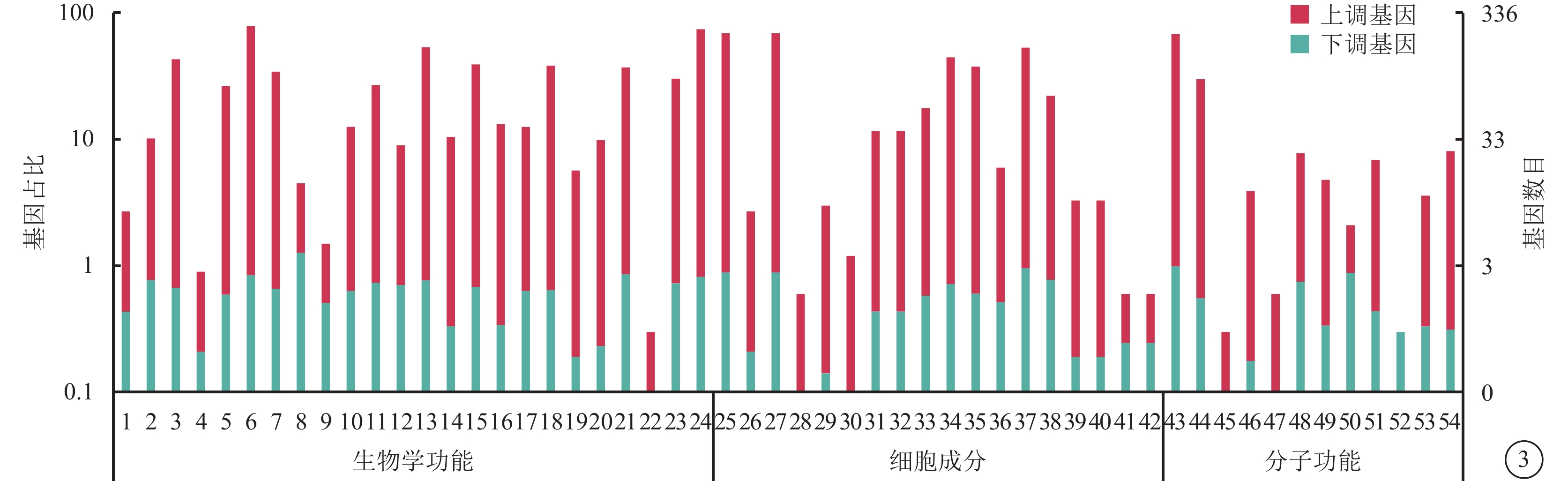
對照組和高糖組之間共有449個差異表達基因。其中,上調基因297個,下調基因152個(圖1)。Pathway顯著性富集分析前20條具有明顯差異的通路,發現差異表達比較明顯的是轉化生長因子-β(TGF-β)信號通路、補體通路等,色氨酸、絲氨酸、氰氨酸等與氨基酸代謝相關的通路也受到了影響。其中,白細胞抑制因子9(SMAD9)、骨形成蛋白4(BMP4)通過TGF-β信號通路發揮作用(圖2)。GO功能顯著性富集分析結果顯示,差異基因的功能主要可以劃分為生物學行為調控,細胞組分形成以及分子功能三大板塊,主要集中表現在生物學過程部分的系統形成和調節多細胞有機體的形成等(圖3)。
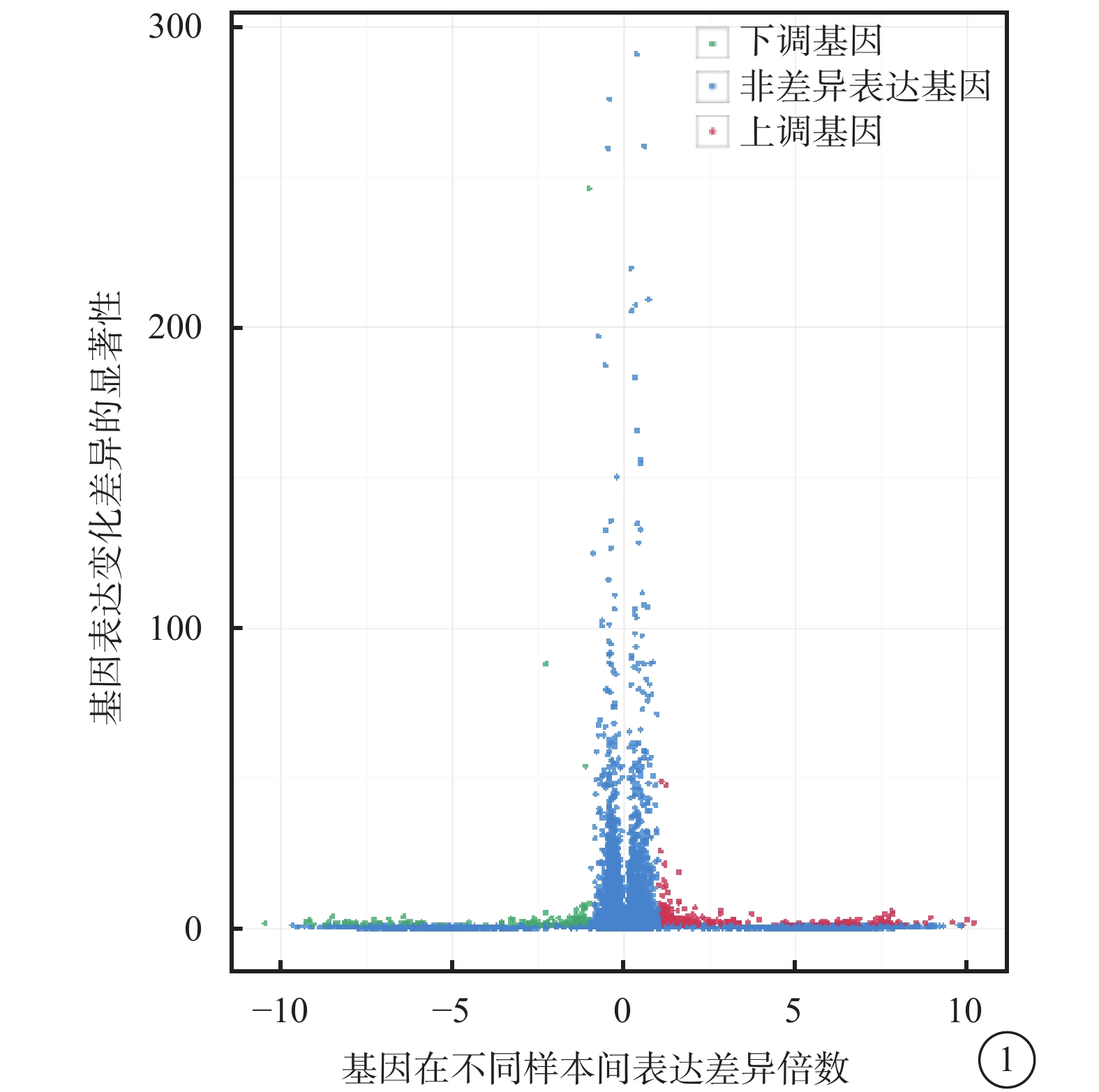 圖1
對照組及高糖組差異表達基因火山圖
圖1
對照組及高糖組差異表達基因火山圖
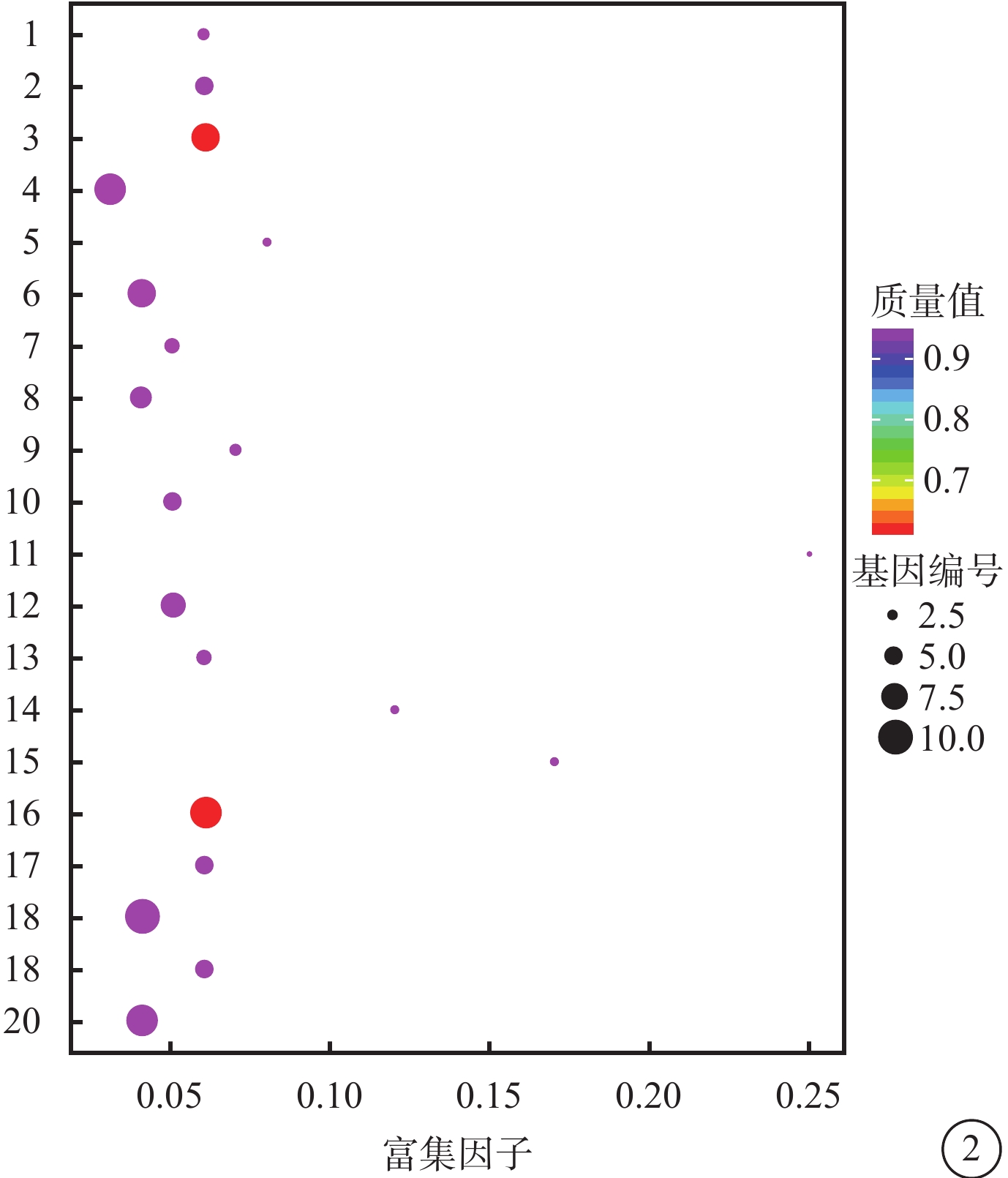 圖2
富集程度排名前20的Pathway條目統計散點圖 1. 維生素的消化和吸收;2. 色氨酸代謝;3. TGF-β信號通路;4. 系統性紅斑狼瘡;5. 類固醇生物合成;6. 金黃色葡萄球菌感染;7. 視黃醇的新陳代謝;8. NOD樣受體信號通路;9. 尼古丁上癮;10. 礦物質的吸收;11. 賴氨酸生物合成;12. 利什曼病;13. 刺猬信號通路;14. 粘多糖生物合成硫酸角質素;15. 氰氨基酸代謝;16. 凝血級聯反應;17. 碳水化合物的消化和吸收;18. 軸突信號;19. 花生四烯酸代謝;20. 阿米巴病
圖2
富集程度排名前20的Pathway條目統計散點圖 1. 維生素的消化和吸收;2. 色氨酸代謝;3. TGF-β信號通路;4. 系統性紅斑狼瘡;5. 類固醇生物合成;6. 金黃色葡萄球菌感染;7. 視黃醇的新陳代謝;8. NOD樣受體信號通路;9. 尼古丁上癮;10. 礦物質的吸收;11. 賴氨酸生物合成;12. 利什曼病;13. 刺猬信號通路;14. 粘多糖生物合成硫酸角質素;15. 氰氨基酸代謝;16. 凝血級聯反應;17. 碳水化合物的消化和吸收;18. 軸突信號;19. 花生四烯酸代謝;20. 阿米巴病
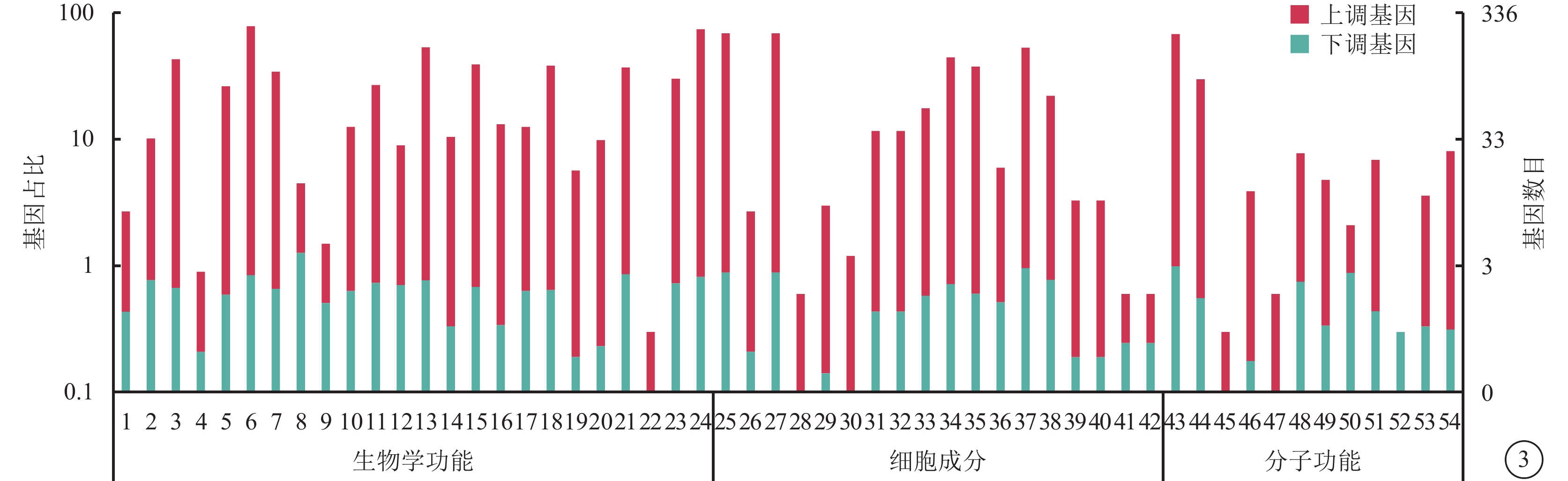 圖3
差異表達基因GO功能注釋分類統計圖 1. 行為;2. 生物粘附;3. 生物調節;4. 細胞死亡;5. 細胞成分組織或生物發生;6. 細胞過程;7. 發展過程;8. 增長;9. 激素分泌;10. 免疫系統的過程;11. 定位;12. 運轉;13. 代謝過程;14. 多生物體合成過程;15. 多細胞有機體合成過程;16. 生物過程的負調節;17. 生物過程的正調節;18. 生物過程的調節;19. 繁殖;20. 繁殖過程;21. 調節反應;22. 節律過程;23. 信號;24. 單生物體合成過程;25. 細胞;26. 細胞連接;27. 細胞成分;28. 膠原蛋白三聚物;29. 細胞外基質;30. 細胞外基質成分;31. 細胞外區域;32. 細胞外區域成分;33. 大分子組合體;34. 膜;35. 膜成分;36. 膜封閉腔;37. 細胞器;38. 細胞器成分;39. 突觸;40. 突觸成分;41. 病毒體;42. 病毒體成分;43. 連接;44. 催化活性;45. 電子載體活動;46. 酶調節活動;47. 鳥苷酸交互獲得;48. 分子傳遞活動;49. 核酸結合轉錄因子活性;50. 蛋白質結合轉錄因子活性;51. 受體活動;52. 受體調節活動;53. 結構分子活動;54. 傳輸活動
圖3
差異表達基因GO功能注釋分類統計圖 1. 行為;2. 生物粘附;3. 生物調節;4. 細胞死亡;5. 細胞成分組織或生物發生;6. 細胞過程;7. 發展過程;8. 增長;9. 激素分泌;10. 免疫系統的過程;11. 定位;12. 運轉;13. 代謝過程;14. 多生物體合成過程;15. 多細胞有機體合成過程;16. 生物過程的負調節;17. 生物過程的正調節;18. 生物過程的調節;19. 繁殖;20. 繁殖過程;21. 調節反應;22. 節律過程;23. 信號;24. 單生物體合成過程;25. 細胞;26. 細胞連接;27. 細胞成分;28. 膠原蛋白三聚物;29. 細胞外基質;30. 細胞外基質成分;31. 細胞外區域;32. 細胞外區域成分;33. 大分子組合體;34. 膜;35. 膜成分;36. 膜封閉腔;37. 細胞器;38. 細胞器成分;39. 突觸;40. 突觸成分;41. 病毒體;42. 病毒體成分;43. 連接;44. 催化活性;45. 電子載體活動;46. 酶調節活動;47. 鳥苷酸交互獲得;48. 分子傳遞活動;49. 核酸結合轉錄因子活性;50. 蛋白質結合轉錄因子活性;51. 受體活動;52. 受體調節活動;53. 結構分子活動;54. 傳輸活動
從449個差異表達基因中,選出差異基因表達較為顯著的11個基因,分析其各自特征及其在對照組與高糖組的表達情況。結果顯示,這些差異表達基因參與分子生物學過程,影響細胞能量代謝、蛋白合成等。其中,ITGB1BP2、UNC5C等與炎癥的產生相關;ATP1A3、CHST5、LCTL等與細胞的能量代謝相關;DAB1、PRSS55等與蛋白合成相關;SMAD9、BMP4等與細胞外基質的代謝相關(表2)。
3 討論
轉錄組研究能夠從整體水平研究基因功能以及基因結構,揭示特定生物學過程以及疾病發生過程中的分子機理,已廣泛應用于基礎研究、臨床診斷和藥物研發等領域。基于Illumina高通量測序平臺的轉錄組測序技術能夠在單核苷酸水平對任意物種的整體轉錄活動進行檢測,在分析轉錄本的結構和表達水平的同時,還能發現未知轉錄本和稀有轉錄本,精確地識別可變剪切位點以及編碼序列單核苷酸多態性,提供最全面的轉錄組信息。相對于傳統的芯片雜交平臺,轉錄組測序無需預先針對已知序列設計探針,即可對任意物種的整體轉錄活動進行檢測,提供更精確的數字化信號、更高的檢測通量以及更廣泛的檢測范圍,是目前深入研究轉錄組復雜性的強大工具。
本研究首次通過RNA轉錄組測序分析來研究高糖狀態對視網膜血管內皮細胞的影響,并發現高糖狀態導致血管內皮細胞基因表達整體化改變,繼而通過GO功能、Pathway顯著性富集分析來探索該樣本中改變的基因其功能及具體的作用通路。結果顯示,高糖對視網膜血管內皮細胞功能改變的過程中涉及到細胞的各種生物學過程改變,由RNA測序得出的449個差異表達基因中,SMAD9、AKR1C4、ITGB1BP2等基因與這種改變密切相關。與此同時也發現許多新的異常表達基因,這些差異表達基因參與分子生物學過程,影響細胞的能量代謝、蛋白合成等。此外,很多與氨基酸代謝相關的通路也受到了影響,如色氨酸、絲氨酸、氰氨酸等代謝通路。氨基酸作為糖尿病代謝網絡中的重要物質,它的代謝異常與糖尿病及其并發癥的發生發展存在著密切關系。
這些差異表達的基因都可通過特定的代謝通路使細胞的功能發生變化,其中差異表達比較明顯的是TGF-β信號通路。我們發現,視網膜血管內皮細胞的TGF-β信號通路在受到高糖誘導后,整體呈上調趨勢,其中多個基因發生明顯上調,如SMAD9、BMP4等。SMAD9可以參與合成TGF-β1信號傳導的蛋白,該類蛋白可以使TGF-β1與其細胞表面絲氨酸蛋白受體結合,進而將TGF-β信號從細胞膜表面受體傳到細胞核,是一種重要的細胞間信號介導分子[7, 8]。BMP4所編碼的蛋白可以與血清中的絲氨酸激酶受體結合,即BMPR1和BMPR2,通過這些受體進行信號傳導,進而通過SMAD通路來影響其目標基因的轉錄。TGF-β1/SMAD信號通路在導致細胞外基質代謝異常以及促進DR發病中起著重要的作用。
近年研究證明,DR是一種慢性持續性炎癥性疾病,各種炎癥因子在DR患者的血清、玻璃體、房水中增加[9, 10]。本研究結果顯示,與炎癥相關的補體系統通路表達上升。補體系統是由補體及其相關因子和膜蛋白組成的機體免疫反應系統。該系統被激活后,可產生清除免疫復合物、介導炎癥反應等效益[11]。補體活化的終末產物C5b-9廣泛沉積在糖尿病患者的視網膜中,補體系統C5是參與DR發生發展的重要因素,其機制與C3a的化學趨化作用有關[12-15]。補體活化時產生的片段導致炎癥細胞聚集并釋放補體C3a,C3a可刺激肥大細胞釋放組胺以及其他活性物質,引起血管擴張、毛細血管壁通透性增加、平滑肌收縮等反應,從而導致視網膜血管內皮細胞損傷[16]。特異性拮抗C3aR可以減輕機體的炎癥反應和組織損傷[17]。我們發現,與炎癥相關的基因表達也呈上調狀態,如NCF1等。NCF1基因參與編碼還原型煙酰胺腺嘌呤二核苷酸磷酸(NADPH)氧化酶的過程。NADPH氧化酶在T細胞介導的炎癥反應中起著重要的作用,NADPH氧化酶活性的降低不僅降低活性氧的產生和病原體清除率,也損害了機體調節炎癥反應的能力[18, 19]。NADPH氧化酶蛋白的表達及激活受高血糖、高血脂、血管緊張素Ⅱ、糖基化終產物等多種因素的影響。Adaikalakoteswari等[20]研究發現,NADPH氧化酶活性與糖尿病病程呈正相關。由此可見,炎癥反應與DR的發生發展密切相關。
從整體上看,高糖對視網膜血管內皮細胞的影響是多層次、多方面的,通過破壞視網膜血管內皮細胞的跨膜傳導、細胞外基質代謝以及蛋白質的轉錄和翻譯等,從而影響視網膜血管內皮細胞的功能。本研究首次通過RNA轉錄組測序分析對高糖作用于視網膜血管內皮細胞的分子作用機制進行了有益的探索,為在分子水平闡述高糖對視網膜血管內皮細胞的作用機制奠定了基礎。至于變化明顯的代謝通路和差異表達基因,有待于今后進一步研究。
研究發現,糖尿病視網膜病變(DR)初期就已出現血管內皮細胞功能異常,且隨著血糖升高,其受損進一步加重[1]。血糖濃度增高可抑制血管舒張,在高糖狀態下,視網膜血管的舒張功能減退,血流速度下降[2]。但鑒于高糖對血管內皮細胞的干預是一個多方面、多層次的復雜過程,我們有必要利用更為先進的檢測方法,在更廣泛的檢測范圍內了解高糖作用下血管內皮細胞改變的完整信息,以期對疾病的后續預防與治療提供有力的依據。RNA轉錄組分析是利用新一代高通量測序技術對全轉錄組測序研究的技術,轉錄組測序的研究對象為特定細胞在某一功能狀態下所能轉錄出來的所有RNA的總和[3]。轉錄組是連接基因組遺傳信息與生物功能蛋白質組的必然紐帶,也是基因功能及結構研究的基礎和出發點。通過新一代高通量測序,能夠全面快速地獲得某一物種特定組織或器官在特定狀態下的幾乎所有轉錄本序列信息,已廣泛應用于基礎研究、臨床診斷和藥物研發等領域[4, 5]。本研究基于RNA轉錄組分析技術,研究高糖狀態對視網膜血管內皮細胞的影響,旨在全面完整的挖掘并分析高糖狀態對血管內皮細胞影響的生物學信息,從而為DR的治療提供更為準確的切入點。
1 材料和方法
1.1 主要材料、細胞培養及實驗分組
恒河猴視網膜血管內皮細胞(RF-6A),由天津醫科大學眼科醫院研究所提供。磷酸鹽緩沖液(PBS)、RPMI-1640培養液(上海源培生物科技股份有限公司),胰蛋白酶、胎牛血清、青鏈霉素、谷氨酰胺(美國Gibco公司)。6孔板、96孔板(美國Life Technologies公司),Trizol試劑盒(美國Invitrogen公司)。葡萄糖(美國Sigma公司)。
RF-6A細胞采用含4.5 ml胎牛血清、0.45 ml雙抗、0.45 ml谷氨酰胺的RPMI-1640培養基于37℃、5%的CO2培養箱中培養,3~5 d傳代1次。取對數生長期細胞用于實驗。實驗分為對照組、高糖組進行,兩組細胞分別采用5、25 mmol/L的葡萄糖持續培養48 h。
1.2 轉錄組文庫測序
轉錄組文庫測序的詳細方法參照文獻[6]。提取不同分組細胞中的總RNA,并對所提取的總RNA的質量進行多方面的質控,以確保后續處理的數據質量合格。隨后在確定總RNA樣品檢測合格后,應用磁珠對poly A尾的mRNA進行富集及片段化,并依據NEB普通建庫方式及鏈特異性建庫方式進行建庫。文庫構建完成后,在經熒光計進行初步定量的基礎之上,應用熒光定量逆轉錄來準確定量文庫有效濃度(文庫有效濃度高于2 nmol/L),以保證文庫質量。庫檢合格后,將不同文庫按照有效濃度及目標下機數據量的需求進行數據匯總分析,經Illumina HiSeq/MiSeq測序儀通過捕獲熒光信號并應用計算機軟件將光信號轉化為測序峰,從而獲得待測片段的序列信息。
1.3 差異表達基因及功能富集分析
將得到的表達譜進行差異表達分析。采用軟件Bioconductor中的edgeR函數做差異表達分析,edgeR函數假設測序reads的計數對于每個基因來說是負二項分布,基于此理論分布來做假設檢驗。并將差異顯著的基因(P≤0.001)提取相應的蛋白質序列進行GO功能、Pathway顯著性富集分析。
應用Blast軟件比對基因序列到NR庫和KEGG數據庫,隨后提取所有基因的GO、Pathway注釋信息。根據差異表達基因列表,應用軟件Bioconductor中的topGO函數和超幾何檢驗對差異表達基因進行Pathway、GO功能顯著性富集分析。對得到的差異基因大數據進行篩查,篩除Log-2到Log2區間內無意義的數據,將該數據按P值大小逐一從數據庫中找出對應基因,查閱相關文獻,了解該基因在不同組織中發揮的作用及作用機制,根據基因的作用途徑,完善顯著差異基因間的關系。
2 結果
所有樣品在illumina測序平臺的轉錄組測序后,得到對照組和高糖組總的reads數分別有50 757 454和49 120 408條;過濾質量較低的數據,得到clean reads數所占比例分別為94.30%、95.14%(表1)。
對照組和高糖組之間共有449個差異表達基因。其中,上調基因297個,下調基因152個(圖1)。Pathway顯著性富集分析前20條具有明顯差異的通路,發現差異表達比較明顯的是轉化生長因子-β(TGF-β)信號通路、補體通路等,色氨酸、絲氨酸、氰氨酸等與氨基酸代謝相關的通路也受到了影響。其中,白細胞抑制因子9(SMAD9)、骨形成蛋白4(BMP4)通過TGF-β信號通路發揮作用(圖2)。GO功能顯著性富集分析結果顯示,差異基因的功能主要可以劃分為生物學行為調控,細胞組分形成以及分子功能三大板塊,主要集中表現在生物學過程部分的系統形成和調節多細胞有機體的形成等(圖3)。
圖1
對照組及高糖組差異表達基因火山圖
圖2
富集程度排名前20的Pathway條目統計散點圖 1. 維生素的消化和吸收;2. 色氨酸代謝;3. TGF-β信號通路;4. 系統性紅斑狼瘡;5. 類固醇生物合成;6. 金黃色葡萄球菌感染;7. 視黃醇的新陳代謝;8. NOD樣受體信號通路;9. 尼古丁上癮;10. 礦物質的吸收;11. 賴氨酸生物合成;12. 利什曼病;13. 刺猬信號通路;14. 粘多糖生物合成硫酸角質素;15. 氰氨基酸代謝;16. 凝血級聯反應;17. 碳水化合物的消化和吸收;18. 軸突信號;19. 花生四烯酸代謝;20. 阿米巴病
圖3
差異表達基因GO功能注釋分類統計圖 1. 行為;2. 生物粘附;3. 生物調節;4. 細胞死亡;5. 細胞成分組織或生物發生;6. 細胞過程;7. 發展過程;8. 增長;9. 激素分泌;10. 免疫系統的過程;11. 定位;12. 運轉;13. 代謝過程;14. 多生物體合成過程;15. 多細胞有機體合成過程;16. 生物過程的負調節;17. 生物過程的正調節;18. 生物過程的調節;19. 繁殖;20. 繁殖過程;21. 調節反應;22. 節律過程;23. 信號;24. 單生物體合成過程;25. 細胞;26. 細胞連接;27. 細胞成分;28. 膠原蛋白三聚物;29. 細胞外基質;30. 細胞外基質成分;31. 細胞外區域;32. 細胞外區域成分;33. 大分子組合體;34. 膜;35. 膜成分;36. 膜封閉腔;37. 細胞器;38. 細胞器成分;39. 突觸;40. 突觸成分;41. 病毒體;42. 病毒體成分;43. 連接;44. 催化活性;45. 電子載體活動;46. 酶調節活動;47. 鳥苷酸交互獲得;48. 分子傳遞活動;49. 核酸結合轉錄因子活性;50. 蛋白質結合轉錄因子活性;51. 受體活動;52. 受體調節活動;53. 結構分子活動;54. 傳輸活動
從449個差異表達基因中,選出差異基因表達較為顯著的11個基因,分析其各自特征及其在對照組與高糖組的表達情況。結果顯示,這些差異表達基因參與分子生物學過程,影響細胞能量代謝、蛋白合成等。其中,ITGB1BP2、UNC5C等與炎癥的產生相關;ATP1A3、CHST5、LCTL等與細胞的能量代謝相關;DAB1、PRSS55等與蛋白合成相關;SMAD9、BMP4等與細胞外基質的代謝相關(表2)。
3 討論
轉錄組研究能夠從整體水平研究基因功能以及基因結構,揭示特定生物學過程以及疾病發生過程中的分子機理,已廣泛應用于基礎研究、臨床診斷和藥物研發等領域。基于Illumina高通量測序平臺的轉錄組測序技術能夠在單核苷酸水平對任意物種的整體轉錄活動進行檢測,在分析轉錄本的結構和表達水平的同時,還能發現未知轉錄本和稀有轉錄本,精確地識別可變剪切位點以及編碼序列單核苷酸多態性,提供最全面的轉錄組信息。相對于傳統的芯片雜交平臺,轉錄組測序無需預先針對已知序列設計探針,即可對任意物種的整體轉錄活動進行檢測,提供更精確的數字化信號、更高的檢測通量以及更廣泛的檢測范圍,是目前深入研究轉錄組復雜性的強大工具。
本研究首次通過RNA轉錄組測序分析來研究高糖狀態對視網膜血管內皮細胞的影響,并發現高糖狀態導致血管內皮細胞基因表達整體化改變,繼而通過GO功能、Pathway顯著性富集分析來探索該樣本中改變的基因其功能及具體的作用通路。結果顯示,高糖對視網膜血管內皮細胞功能改變的過程中涉及到細胞的各種生物學過程改變,由RNA測序得出的449個差異表達基因中,SMAD9、AKR1C4、ITGB1BP2等基因與這種改變密切相關。與此同時也發現許多新的異常表達基因,這些差異表達基因參與分子生物學過程,影響細胞的能量代謝、蛋白合成等。此外,很多與氨基酸代謝相關的通路也受到了影響,如色氨酸、絲氨酸、氰氨酸等代謝通路。氨基酸作為糖尿病代謝網絡中的重要物質,它的代謝異常與糖尿病及其并發癥的發生發展存在著密切關系。
這些差異表達的基因都可通過特定的代謝通路使細胞的功能發生變化,其中差異表達比較明顯的是TGF-β信號通路。我們發現,視網膜血管內皮細胞的TGF-β信號通路在受到高糖誘導后,整體呈上調趨勢,其中多個基因發生明顯上調,如SMAD9、BMP4等。SMAD9可以參與合成TGF-β1信號傳導的蛋白,該類蛋白可以使TGF-β1與其細胞表面絲氨酸蛋白受體結合,進而將TGF-β信號從細胞膜表面受體傳到細胞核,是一種重要的細胞間信號介導分子[7, 8]。BMP4所編碼的蛋白可以與血清中的絲氨酸激酶受體結合,即BMPR1和BMPR2,通過這些受體進行信號傳導,進而通過SMAD通路來影響其目標基因的轉錄。TGF-β1/SMAD信號通路在導致細胞外基質代謝異常以及促進DR發病中起著重要的作用。
近年研究證明,DR是一種慢性持續性炎癥性疾病,各種炎癥因子在DR患者的血清、玻璃體、房水中增加[9, 10]。本研究結果顯示,與炎癥相關的補體系統通路表達上升。補體系統是由補體及其相關因子和膜蛋白組成的機體免疫反應系統。該系統被激活后,可產生清除免疫復合物、介導炎癥反應等效益[11]。補體活化的終末產物C5b-9廣泛沉積在糖尿病患者的視網膜中,補體系統C5是參與DR發生發展的重要因素,其機制與C3a的化學趨化作用有關[12-15]。補體活化時產生的片段導致炎癥細胞聚集并釋放補體C3a,C3a可刺激肥大細胞釋放組胺以及其他活性物質,引起血管擴張、毛細血管壁通透性增加、平滑肌收縮等反應,從而導致視網膜血管內皮細胞損傷[16]。特異性拮抗C3aR可以減輕機體的炎癥反應和組織損傷[17]。我們發現,與炎癥相關的基因表達也呈上調狀態,如NCF1等。NCF1基因參與編碼還原型煙酰胺腺嘌呤二核苷酸磷酸(NADPH)氧化酶的過程。NADPH氧化酶在T細胞介導的炎癥反應中起著重要的作用,NADPH氧化酶活性的降低不僅降低活性氧的產生和病原體清除率,也損害了機體調節炎癥反應的能力[18, 19]。NADPH氧化酶蛋白的表達及激活受高血糖、高血脂、血管緊張素Ⅱ、糖基化終產物等多種因素的影響。Adaikalakoteswari等[20]研究發現,NADPH氧化酶活性與糖尿病病程呈正相關。由此可見,炎癥反應與DR的發生發展密切相關。
從整體上看,高糖對視網膜血管內皮細胞的影響是多層次、多方面的,通過破壞視網膜血管內皮細胞的跨膜傳導、細胞外基質代謝以及蛋白質的轉錄和翻譯等,從而影響視網膜血管內皮細胞的功能。本研究首次通過RNA轉錄組測序分析對高糖作用于視網膜血管內皮細胞的分子作用機制進行了有益的探索,為在分子水平闡述高糖對視網膜血管內皮細胞的作用機制奠定了基礎。至于變化明顯的代謝通路和差異表達基因,有待于今后進一步研究。