引用本文: 周賢慧, 孟旭霞, 胡迭. 軸突導向因子-1對糖尿病大鼠視網膜Müller細胞活化的影響. 中華眼底病雜志, 2017, 33(5): 527-529. doi: 10.3760/cma.j.issn.1005-1015.2017.05.020 復制
研究發現,糖尿病(DM)視網膜病變(DR)在視網膜血管發生改變之前其視網膜神經膠質細胞即出現異常[1]。Müller細胞是視網膜最主要的膠質細胞。在DR進程中,異常活化的Müller細胞促進血管內皮生長因子(VEGF)及堿性成纖維細胞生長因子釋放,導致視網膜新生血管生成及纖維化[2, 3]。軸突導向因子(netrin-1)是目前已知的最主要神經生長導向因子。近年研究發現,netrin-1在新生血管生成中發揮重要作用[4, 5]。我們推測,如能通過外源性netrin-1早期干預Müller細胞,降低其異常活化,減少視網膜新生血管形成,可能減緩DR進程。為驗證這一推測,本研究觀察了外源性netrin-1對DM大鼠視網膜Müller細胞活化的影響。現將結果報道如下。
1 材料和方法
健康清潔級成年雄性Sprague-Dawley大鼠50只,體重約250 g,由山東魯抗醫藥公司提供。所有動物的飼養及管理均符合國家科學技術委員會頒布的《實驗動物管理條例》。采用隨機數字表法將50只大鼠隨機分為正常對照組(A組)、正常+平衡鹽溶液(BSS)組(B組)、正常+netrin-1 100 μg/ml組(C組)、DM+BSS組(D組)、DM+netrin-1 100 μg/ml組(E組),每組10只。D、E組大鼠按60 mg/kg的劑量,左下腹腔注射鏈脲佐菌素誘導DM動物模型。以注射72 h后空腹血糖≥16.5 mmol/L為DM模型建立成功[6]。建模成功率為100%。建模3個月后,C、E組大鼠玻璃體腔分別注射100 μg/ml netrin-1 5 μl。B、D組大鼠玻璃體腔注射等體積BSS。A組大鼠不做任何處理。
10%水合氯醛按0.3 ml/100 g劑量腹腔注射麻醉大鼠,行前房穿刺,放出少量房水,微量加樣器取5 μl netrin-1自大鼠顳上方角膜緣后1.5~2.0 mm處以45°角刺入玻璃體腔,緩慢注入netrin-1,注射完畢后按壓穿刺口3 min,無滲出,涂氧氟沙星眼膏。所有實驗大鼠均未出現注射相關的高眼壓及鞏膜口漏等并發癥。
采用免疫組織化學染色法檢測大鼠視網膜膠質原纖維酸性蛋白(GFAP)的表達。玻璃體腔注射netrin-1后1個月,過量麻醉處死大鼠,迅速摘除眼球作石蠟切片。常規脫蠟和水化后,5%山羊血清封閉,一抗用1:200兔抗大鼠GFAP多克隆抗體,二抗用生物素標記山羊抗兔IgG抗體,二氨基聯苯胺(DAB)顯色,蘇木精復染,中性樹膠封片。光學顯微鏡下觀察并照相,以血管內皮細胞膜呈棕黃色為陽性表達。采用Image pro plus 6.0軟件對圖像進行分析。以平均積分吸光度[A,舊稱光密度(OD)]值表示視網膜中GFAP的表達量。
采用熒光定量聚合酶鏈反應檢測大鼠視網膜GFAP mRNA表達。玻璃體腔注射netrin-1后1個月,過量麻醉處死大鼠。視網膜稱重,按照每50~l00 mg加1 ml RNAiso Plus裂解視網膜,提取視網膜總RNA,去除RNA內殘留的DNA,逆轉錄成cDNA,以β-肌動蛋白(β-actin)作為內參照。用GFAP引物SYBR GreenⅠ嵌合熒光法擴增相應基因。GFAP:上游引物5′-ACCGCATCACCATTCCTGTA-3′,下游引物5′-GCACACCTCACATCACATCC-3′。β-actin:上游引物5′-TGGCACCCAGCACAATGAA-3′,下游引物5′-CTAAGTCATAGTCCGCCTAGAAGCA-3′。退火溫度60℃,40個循環后形成擴增曲線,記錄熒光值到達閾值時所經歷的循環閾值。將A組大鼠視網膜GFAP mRNA表達設定為1,計算其他各組大鼠視網膜GFAP mRNA表達。
采用SPSS16.0統計軟件進行統計學分析。定量資料結果以均數±標準差(
 )表示,兩獨立樣本間比較采用獨立樣本 t 檢驗,多組間比較采用單因素方差分析。P<0.05為差異有統計學意義。
)表示,兩獨立樣本間比較采用獨立樣本 t 檢驗,多組間比較采用單因素方差分析。P<0.05為差異有統計學意義。
2 結果
免疫組織化學染色結果顯示,A~C組大鼠視網膜僅在神經纖維層及神經節細胞層可見少量GFAP陽性表達。D組大鼠視網膜GFAP陽性表達增強。E組大鼠視網膜GFAP陽性表達較D組減少(圖1)。
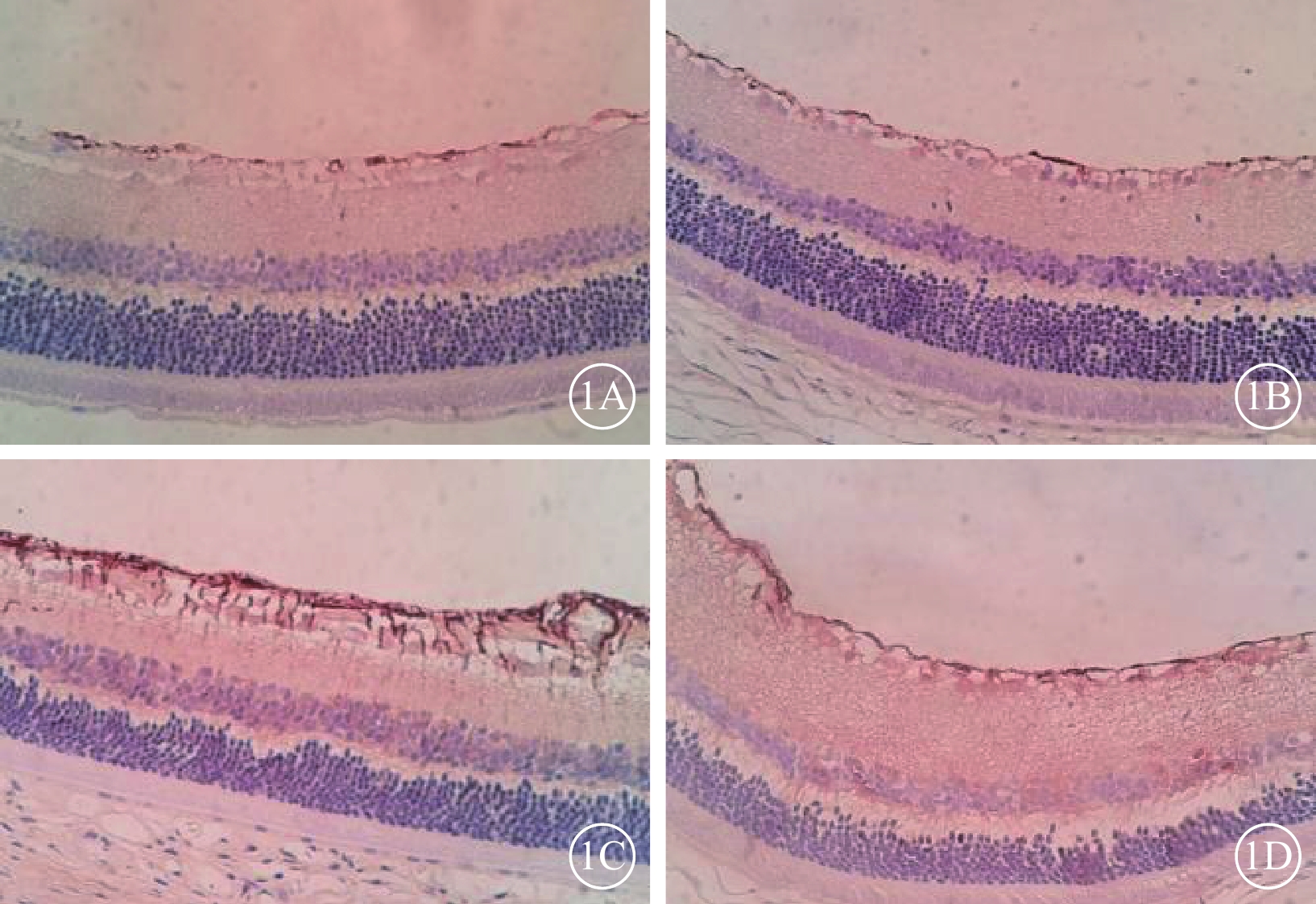 圖1
大鼠視網膜GFAP免疫組織化學染色像。1A. B組;1B. C組;1C. D組;1D. E組。B、C組大鼠視網膜僅在神經纖維層及神經節細胞層可見少量GFAP陽性表達;D組大鼠視網膜GFAP陽性表達增多;E組大鼠視網膜GFAP陽性表達較D組減少 DAB ×40
圖1
大鼠視網膜GFAP免疫組織化學染色像。1A. B組;1B. C組;1C. D組;1D. E組。B、C組大鼠視網膜僅在神經纖維層及神經節細胞層可見少量GFAP陽性表達;D組大鼠視網膜GFAP陽性表達增多;E組大鼠視網膜GFAP陽性表達較D組減少 DAB ×40
A~E組大鼠視網膜GFAP陽性表達、mRNA表達比較,差異有統計學意義(F=203.43、72.91,P=0.00、0.00)(表1)。與A組比較,D組大鼠視網膜GFAP陽性表達、mRNA表達明顯增多,差異有統計學意義(t=?26.01、22.26,P=0.00、0.00)。E組大鼠視網膜GFAP陽性表達、mRNA表達較D組明顯下降(t=?10.78、3.93,P=0.00、0.00);但仍高于A組(t=7.00、?9.82,P=0.00、0.00),差異均有統計學意義。A、C組大鼠視網膜GFAP陽性表達、mRNA表達比較,差異均無統計學意義(t=?0.29、0.50,P=0.77、0.62)。
 )
)
3 討論
Müller細胞是視網膜最主要的膠質細胞,位于視網膜內界膜與外界膜之間,橫跨整個視網膜;參與構成血視網膜屏障,支撐視網膜結構,釋放神經遞質和營養因子,保護及營養視神經細胞等重要作用[7]。GFAP是膠質細胞的特異性標記物。正常大鼠Müller細胞極少甚至不表達GFAP;而在DR中,Müller細胞異常活化,分泌生長因子和炎性介質,導致視網膜神經細胞和毛細血管內皮細胞功能障礙、周細胞凋亡,促進視網膜新生血管形成[8, 9]。Fu等[10]發現,Müller細胞在DM視神經細胞存活過程中也發揮重要作用。因此,保護DM大鼠Müller細胞,減少其異常活化顯得尤為重要。
Netrin-1是目前已知的最主要神經導向因子,介導細胞遷移、黏附、分化和新生血管生成等作用[4, 5]。DR時,視網膜膠質細胞異常活化神經元細胞凋亡[11],視網膜血管通透性改變,新生血管生成;而netrin-1具有介導突觸生長,保護神經元及降低視網膜血管通透性的作用[12]。我們不禁假設netrin-1是否在兩者之間存在某種共通性,是否在DR中起到“橋梁”作用,是否對膠質細胞的活化有影響,而Müller細胞作為主要的膠質細胞成為觀察焦點。本研究結果顯示,D、E組大鼠Müller細胞活化明顯增加,其標志物GFAP陽性表達增多,GFAP mRNA隨之升高;當給予外源性netrin-1干預時,視網膜GFAP表達降低,Müller細胞活性下降,但仍未能恢復至正常水平。而A、C組大鼠視網膜GFAP表達之間無明顯差異。據此我們推測,100 μg/ml的外源性netrin-1可降低DM大鼠Müller細胞的活化,減慢DR進程,但是不能阻止DR進展。此外,我們還發現外源性netrin-1對正常大鼠Müller細胞的活化無明顯影響。
本研究結果表明,外源性netrin-1可降低DM大鼠Müller細胞的活化,但其具體機制及信號轉導通路目前尚不明確。我們前期研究發現,netrin-1可降低DM大鼠視網膜血管通透性[16]。但具體是netrin-1通過保護Müller細胞來降低視網膜血管通透性,抑或是通過降低視網膜血管通透性打破神經-血管環路的惡性循環來降低Müller細胞的活性,目前仍不清楚。這需要我們在分子生物學水平對Müller細胞進行干預以及進一步的實驗數據支持。此外,本研究尚未對神經元細胞凋亡進行觀察,這將是我們下一步的研究重點。
志謝 感謝煙臺市業達醫院眼科姜文科、劉曉玲醫師對文稿撰寫提供的幫助
研究發現,糖尿病(DM)視網膜病變(DR)在視網膜血管發生改變之前其視網膜神經膠質細胞即出現異常[1]。Müller細胞是視網膜最主要的膠質細胞。在DR進程中,異常活化的Müller細胞促進血管內皮生長因子(VEGF)及堿性成纖維細胞生長因子釋放,導致視網膜新生血管生成及纖維化[2, 3]。軸突導向因子(netrin-1)是目前已知的最主要神經生長導向因子。近年研究發現,netrin-1在新生血管生成中發揮重要作用[4, 5]。我們推測,如能通過外源性netrin-1早期干預Müller細胞,降低其異常活化,減少視網膜新生血管形成,可能減緩DR進程。為驗證這一推測,本研究觀察了外源性netrin-1對DM大鼠視網膜Müller細胞活化的影響。現將結果報道如下。
1 材料和方法
健康清潔級成年雄性Sprague-Dawley大鼠50只,體重約250 g,由山東魯抗醫藥公司提供。所有動物的飼養及管理均符合國家科學技術委員會頒布的《實驗動物管理條例》。采用隨機數字表法將50只大鼠隨機分為正常對照組(A組)、正常+平衡鹽溶液(BSS)組(B組)、正常+netrin-1 100 μg/ml組(C組)、DM+BSS組(D組)、DM+netrin-1 100 μg/ml組(E組),每組10只。D、E組大鼠按60 mg/kg的劑量,左下腹腔注射鏈脲佐菌素誘導DM動物模型。以注射72 h后空腹血糖≥16.5 mmol/L為DM模型建立成功[6]。建模成功率為100%。建模3個月后,C、E組大鼠玻璃體腔分別注射100 μg/ml netrin-1 5 μl。B、D組大鼠玻璃體腔注射等體積BSS。A組大鼠不做任何處理。
10%水合氯醛按0.3 ml/100 g劑量腹腔注射麻醉大鼠,行前房穿刺,放出少量房水,微量加樣器取5 μl netrin-1自大鼠顳上方角膜緣后1.5~2.0 mm處以45°角刺入玻璃體腔,緩慢注入netrin-1,注射完畢后按壓穿刺口3 min,無滲出,涂氧氟沙星眼膏。所有實驗大鼠均未出現注射相關的高眼壓及鞏膜口漏等并發癥。
采用免疫組織化學染色法檢測大鼠視網膜膠質原纖維酸性蛋白(GFAP)的表達。玻璃體腔注射netrin-1后1個月,過量麻醉處死大鼠,迅速摘除眼球作石蠟切片。常規脫蠟和水化后,5%山羊血清封閉,一抗用1:200兔抗大鼠GFAP多克隆抗體,二抗用生物素標記山羊抗兔IgG抗體,二氨基聯苯胺(DAB)顯色,蘇木精復染,中性樹膠封片。光學顯微鏡下觀察并照相,以血管內皮細胞膜呈棕黃色為陽性表達。采用Image pro plus 6.0軟件對圖像進行分析。以平均積分吸光度[A,舊稱光密度(OD)]值表示視網膜中GFAP的表達量。
采用熒光定量聚合酶鏈反應檢測大鼠視網膜GFAP mRNA表達。玻璃體腔注射netrin-1后1個月,過量麻醉處死大鼠。視網膜稱重,按照每50~l00 mg加1 ml RNAiso Plus裂解視網膜,提取視網膜總RNA,去除RNA內殘留的DNA,逆轉錄成cDNA,以β-肌動蛋白(β-actin)作為內參照。用GFAP引物SYBR GreenⅠ嵌合熒光法擴增相應基因。GFAP:上游引物5′-ACCGCATCACCATTCCTGTA-3′,下游引物5′-GCACACCTCACATCACATCC-3′。β-actin:上游引物5′-TGGCACCCAGCACAATGAA-3′,下游引物5′-CTAAGTCATAGTCCGCCTAGAAGCA-3′。退火溫度60℃,40個循環后形成擴增曲線,記錄熒光值到達閾值時所經歷的循環閾值。將A組大鼠視網膜GFAP mRNA表達設定為1,計算其他各組大鼠視網膜GFAP mRNA表達。
采用SPSS16.0統計軟件進行統計學分析。定量資料結果以均數±標準差(
)表示,兩獨立樣本間比較采用獨立樣本 t 檢驗,多組間比較采用單因素方差分析。P<0.05為差異有統計學意義。
2 結果
免疫組織化學染色結果顯示,A~C組大鼠視網膜僅在神經纖維層及神經節細胞層可見少量GFAP陽性表達。D組大鼠視網膜GFAP陽性表達增強。E組大鼠視網膜GFAP陽性表達較D組減少(圖1)。
圖1
大鼠視網膜GFAP免疫組織化學染色像。1A. B組;1B. C組;1C. D組;1D. E組。B、C組大鼠視網膜僅在神經纖維層及神經節細胞層可見少量GFAP陽性表達;D組大鼠視網膜GFAP陽性表達增多;E組大鼠視網膜GFAP陽性表達較D組減少 DAB ×40
A~E組大鼠視網膜GFAP陽性表達、mRNA表達比較,差異有統計學意義(F=203.43、72.91,P=0.00、0.00)(表1)。與A組比較,D組大鼠視網膜GFAP陽性表達、mRNA表達明顯增多,差異有統計學意義(t=?26.01、22.26,P=0.00、0.00)。E組大鼠視網膜GFAP陽性表達、mRNA表達較D組明顯下降(t=?10.78、3.93,P=0.00、0.00);但仍高于A組(t=7.00、?9.82,P=0.00、0.00),差異均有統計學意義。A、C組大鼠視網膜GFAP陽性表達、mRNA表達比較,差異均無統計學意義(t=?0.29、0.50,P=0.77、0.62)。
)
3 討論
Müller細胞是視網膜最主要的膠質細胞,位于視網膜內界膜與外界膜之間,橫跨整個視網膜;參與構成血視網膜屏障,支撐視網膜結構,釋放神經遞質和營養因子,保護及營養視神經細胞等重要作用[7]。GFAP是膠質細胞的特異性標記物。正常大鼠Müller細胞極少甚至不表達GFAP;而在DR中,Müller細胞異常活化,分泌生長因子和炎性介質,導致視網膜神經細胞和毛細血管內皮細胞功能障礙、周細胞凋亡,促進視網膜新生血管形成[8, 9]。Fu等[10]發現,Müller細胞在DM視神經細胞存活過程中也發揮重要作用。因此,保護DM大鼠Müller細胞,減少其異常活化顯得尤為重要。
Netrin-1是目前已知的最主要神經導向因子,介導細胞遷移、黏附、分化和新生血管生成等作用[4, 5]。DR時,視網膜膠質細胞異常活化神經元細胞凋亡[11],視網膜血管通透性改變,新生血管生成;而netrin-1具有介導突觸生長,保護神經元及降低視網膜血管通透性的作用[12]。我們不禁假設netrin-1是否在兩者之間存在某種共通性,是否在DR中起到“橋梁”作用,是否對膠質細胞的活化有影響,而Müller細胞作為主要的膠質細胞成為觀察焦點。本研究結果顯示,D、E組大鼠Müller細胞活化明顯增加,其標志物GFAP陽性表達增多,GFAP mRNA隨之升高;當給予外源性netrin-1干預時,視網膜GFAP表達降低,Müller細胞活性下降,但仍未能恢復至正常水平。而A、C組大鼠視網膜GFAP表達之間無明顯差異。據此我們推測,100 μg/ml的外源性netrin-1可降低DM大鼠Müller細胞的活化,減慢DR進程,但是不能阻止DR進展。此外,我們還發現外源性netrin-1對正常大鼠Müller細胞的活化無明顯影響。
本研究結果表明,外源性netrin-1可降低DM大鼠Müller細胞的活化,但其具體機制及信號轉導通路目前尚不明確。我們前期研究發現,netrin-1可降低DM大鼠視網膜血管通透性[16]。但具體是netrin-1通過保護Müller細胞來降低視網膜血管通透性,抑或是通過降低視網膜血管通透性打破神經-血管環路的惡性循環來降低Müller細胞的活性,目前仍不清楚。這需要我們在分子生物學水平對Müller細胞進行干預以及進一步的實驗數據支持。此外,本研究尚未對神經元細胞凋亡進行觀察,這將是我們下一步的研究重點。
志謝 感謝煙臺市業達醫院眼科姜文科、劉曉玲醫師對文稿撰寫提供的幫助