引用本文: 郭慶敏, 孟旭霞, 劉鵬輝, 周賢慧. Notch受體蛋白1、Delta樣配體4在增生型糖尿病視網膜病變新生血管形成中的作用. 中華眼底病雜志, 2017, 33(3): 275-280. doi: 10.3760/cma.j.issn.1005-1015.2017.03.013 復制
Notch信號通路在血管生成過程中發揮重要作用[1]。除了維持視網膜正常生理微環境,其缺失將導致視網膜血管和細胞發育不良及視網膜環境穩態失調[2],并且參與缺氧條件下視網膜新生血管的發生[3]。作為Notch信號通路的配體之一,Delta樣配體(Dll)4已被證實可特異性表達于生理及病理新生血管的發生中[4]。我們前期動物模型研究發現,Notch受體蛋白1(Notch1)、Dll4的表達量在早期糖尿病視網膜病變(DR)發展過程中逐漸升高,血管內皮生長因子(VEGF)受體2(VEGFR2)表達量逐漸增加,且VEGFR2的表達與Notch1、Dll4呈正相關[5]。但Notch信號通路是否參與人類增生型DR(PDR)新生血管的發生尚不清楚。因此,我們通過檢測PDR患眼纖維血管膜病理標本中Notch1、Dll4、VEGFR2的表達,進一步探討Notch1、Dll4在PDR新生血管形成中的作用及與VEGF信號通路的關系。現將結果報道如下。
1 對象和方法
2014年7月至2015年7月在青島大學附屬醫院眼科行睫狀體平坦部23G標準三通道玻璃體切割手術的PDR患者57例60只眼納入研究。本研究獲得青島大學附屬醫院倫理審查委員會審查批準;納入研究對象或家屬均簽署知情同意書。患者中男性32例34只眼,女性25例26只眼。年齡36~73歲,平均年齡(54.93±6.42)歲。糖尿病病程8~20年,平均糖尿病病程(15.72±3.51)年。所有患者均符合2型糖尿病診斷標準及PDR診斷標準[6,7]。排除肺部疾病、嚴重肝腎功能不全、心腦血管病變、惡性腫瘤及自身免疫性疾病者。
根據手術前是否行抗VEGF藥物玻璃體腔注射將患眼分為PDR未注藥組和PDR注藥組,分別為30例32只眼、27例28只眼。PDR未注藥組30例32只眼中,男性16例18只眼,女性14例14只眼;年齡36~73歲,平均年齡55歲;糖尿病病程8~20年,平均糖尿病病程16年。PDR注藥組27例28只眼中,男性16例16只眼,女性11例12只眼;年齡40~68歲,平均年齡54歲;糖尿病病程8~18年,平均糖尿病病程15年。玻璃體切割手術前2~7 d行雷珠單抗玻璃體腔注射治療。選取特發性黃斑前膜患者18例18只眼作為對照組。其中,男性5例5只眼,女性13例13只眼;年齡43~79歲,平均年齡61歲。均無糖尿病及糖尿病家族史,眼部及全身感染、急性炎癥反應及缺血缺氧性疾病史以及全身系統性疾病。3組患者年齡(F=1.14)、性別構成比(F=0.97)比較,差異均無統計學意義(P>0.05)。
玻璃體切割手術中應用視網膜剪及視網膜鑷獲得PDR纖維血管膜病理標本及黃斑前膜病理標本。隨機數字表法隨機分配標本。30只眼病理標本立即用4%多聚甲醛溶液固定,常規酒精脫水,二甲苯透明,浸蠟包埋,3 μm連續切片并編號,行光學顯微鏡觀察及蛋白表達檢測;48只眼病理標本則移入已消毒的離心管中,–80 ℃超低溫冰箱中凍存,用于基因檢測。行光學顯微鏡觀察及蛋白表達檢測的30只眼病理標本中,PDR未注藥組、PDR注藥組、對照組分別為10、12、8只眼;行基因檢測的48只眼病理標本中,PDR未注藥組、PDR注藥組、對照組分別為22、16、10只眼。
所有石蠟標本截面最大處連續切片5張進行蘇木精-伊紅(HE)染色。高倍光學顯微鏡下觀察。每張切片隨機選取5個視野采用盲法計數突破內界膜的血管內皮細胞,各切片內皮細胞計數為5個視野中細胞計數的平均值,最后計算各組病理標本每張切片的血管內皮細胞計數的平均值。
免疫組織化學方法檢測各組病理標本中Notch1、Dll4、VEGFR2蛋白表達。每個病理標本選取截面最大處連續切片15張,各選取5張按照免疫組織化學試劑盒說明要求進行Notch1、Dll4及VEGFR2蛋白的免疫組織化學染色,二氨基苯聯胺(DAB)顯色3 min,磷酸鹽緩沖液代替一抗作為陰性對照。結果以細胞漿內出現黃色、棕黃色或棕褐色顆粒為弱陽性、陽性、強陽性。采用Image pro plus6.0軟件計算每張切片相應蛋白免疫組織化學染色陽性部分的平均吸光度[A,舊稱光密度(OD)]值,最后統計各組病理標本中各種蛋白的平均值,代表相應蛋白的表達量。
熒光定量聚合酶鏈反應(RT-PCR)檢測各組病理標本中Notch1、Dll4、VEGFR2 mRNA的表達。50~100 mg纖維血管膜或黃斑前膜加入1 ml RNAiso plus進行裂解;提取總RNA,紫外分光光度計檢測其濃度和純度,A260/A280在1.7~2.1的樣本用于試驗。逆轉錄成cDNA,以磷酸甘油醛脫氫酶(GAPDH)作為內參基因,采用雙鏈嵌合熒光染色法擴增相應基因。引物序列:Notch1:上游引物5′-GTCAACGCCGTAGAT-GACC-3′,下游引物5′-TTGTTAGCCCCGTTCTTCAG-3′,長度101堿基對(bp);Dll4:上游引物5′-CCC-TGGCAATGTACTTGTGAT-3′,下游引物5′-TGGT-GGGTGCAGTAGTTGAG-3,長度74 bp;VEGFR2:上游引物5′-AGTGATCGGAAATGACACTGGA-3′,下游引物5′-GCACAAAGTGACACGTTGAGAT-3′,長度213 bp;GAPDH:上游引物5′-CACGATGGAGGGG-CCGGACTCATC-3′,下游引物5′-TAAAGACCTCTAT-GCCAACACAGT -3′,長度241 bp。退火溫度60 ℃,40個循環后形成擴增曲線,記錄循環閾值(Ct值),結果采用2–ΔΔCt公式進行分析。
采用SPSS19.0統計學軟件進行統計學分析處理。數據經Kolmogoror-Smimor檢驗證實成正態分布后用均數±標準差( )表示。多組間相關因子表達比較采用Kruskal-Wallis檢驗。兩變量間關系采用Spearman相關性分析。α=0.05。P<0.05為差異有統計學意義。
2 結果
光學顯微鏡觀察發現,PDR纖維血管膜病理標本均可見新生血管生成,但新生血管數量和形態不同。PDR注藥組12只眼纖維血管膜病理標本中可見血管管腔較狹窄,部分管腔出現閉合。PDR未注藥組10只眼纖維血管膜病理標本可見血管管腔擴張,形態較完整。對照組8只眼黃斑前膜病理標本中可見各種細胞散布其中,未見血管結構;未見突破內界膜的血管內皮細胞(圖1)。PDR注藥組、PDR未注藥組纖維血管膜病理標本中突破內界膜的血管內皮細胞數量分別為(17.17±2.48)、(41.50±5.57)個;兩組纖維血管膜病理標本突破內界膜的血管內皮細胞數量比較,差異有統計學意義(t=9.58,P<0.05)。

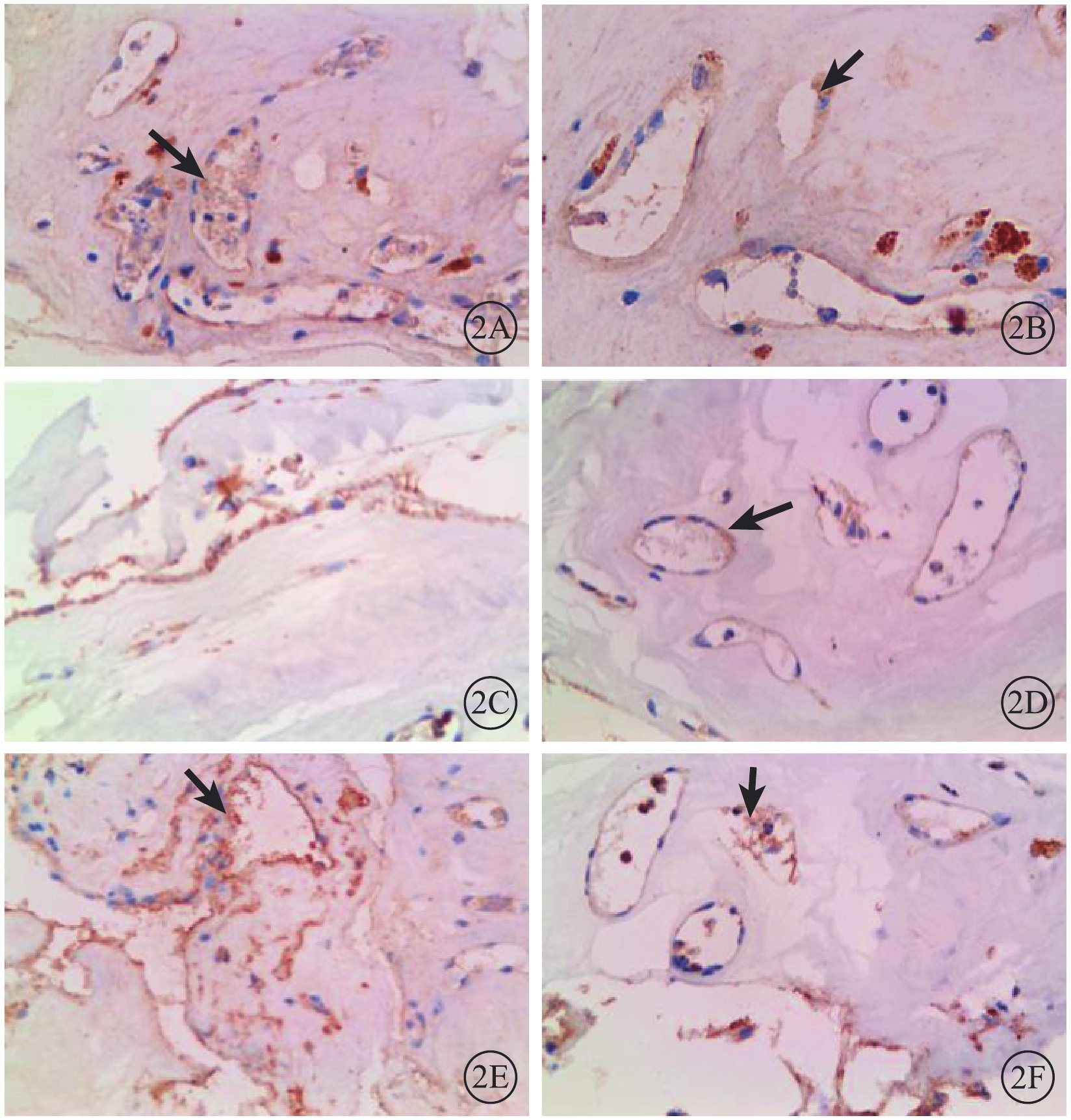
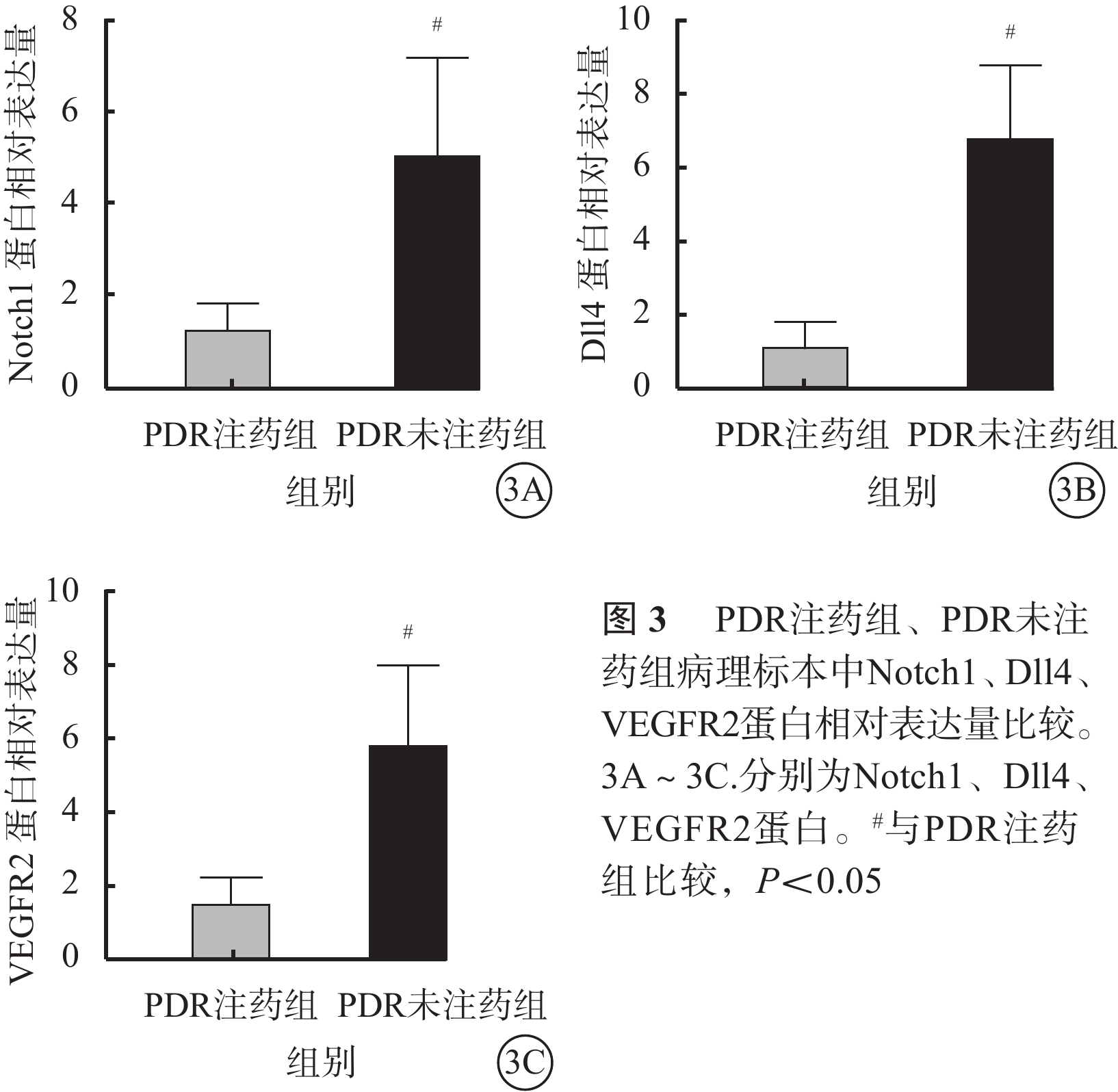
免疫組織化學染色結果顯示,PDR 22只眼纖維血管膜病理標本中Notch1、Dll4、VEGFR2表達均呈陽性,且主要表達在新生血管內皮組織,血管外部位表達較少(圖2)。PDR注藥組、PDR未注藥組纖維血管膜病理標本中Notch1、Dll4、VEGFR2蛋白相對表達量比較,差異均有統計學意義(t=3.45、6.01、4.08,P=0.030、0.008、0.023)(圖3)。
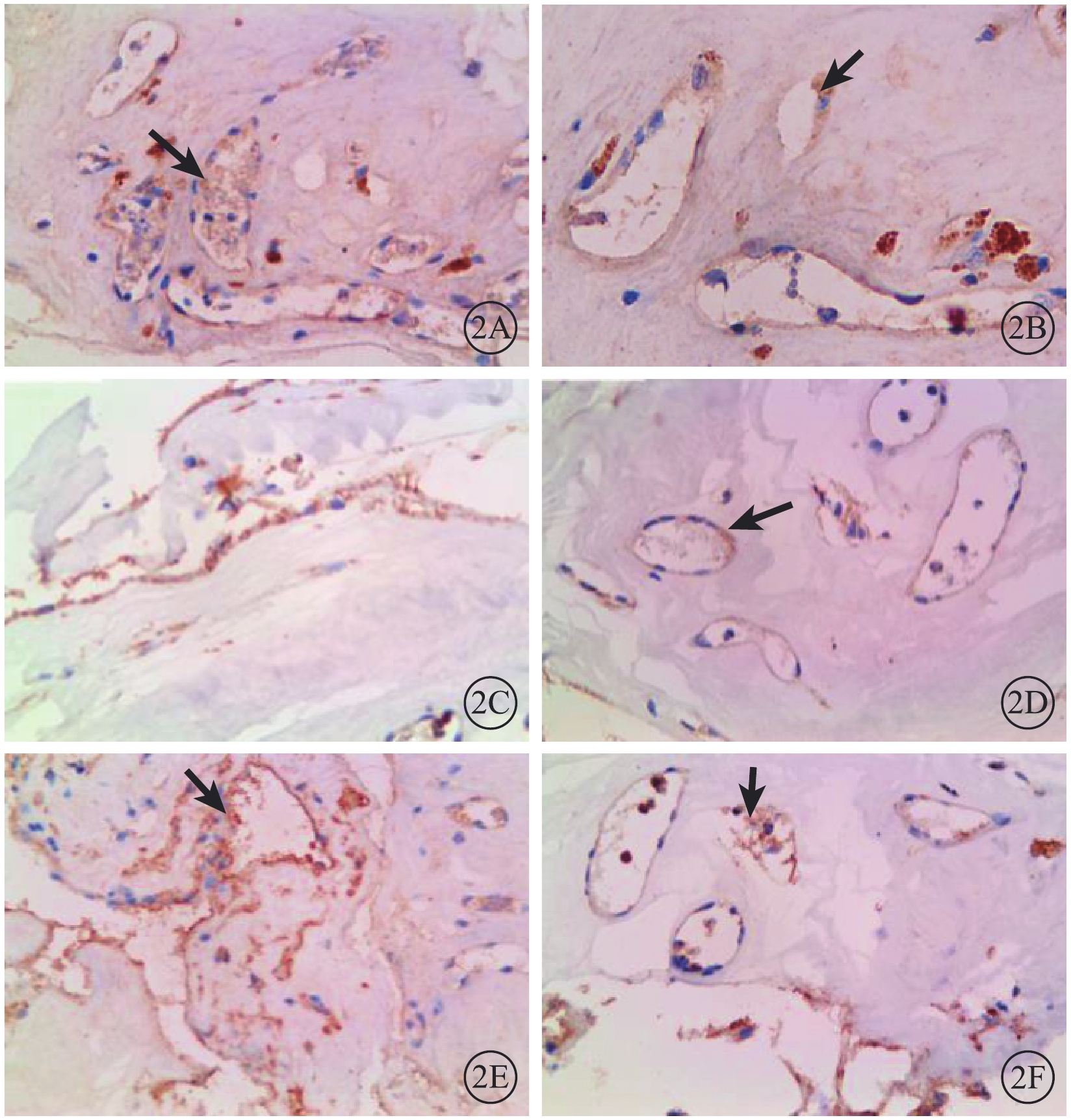 圖2
PDR患眼纖維血管膜病理標本免疫組織化學染色像。2A.PDR未注藥組,Notch1蛋白在新生血管內皮細胞中呈強陽性表達,而在纖維細胞和細胞外基質中幾乎無表達(黑箭) DAB ×400;2B.PDR注藥組,Notch1蛋白在血管內皮細胞中呈弱陽性表達(黑箭) DAB ×400;2C.PDR未注藥組,Dll4蛋白僅在新生血管內皮細胞中呈陽性表達,DAB ×400;2D.PDR注藥組,Dll4蛋白在血管內皮細胞中呈弱陽性表達(黑箭)DAB ×400;2E.PDR未注藥組,VEGFR2蛋白在新生血管內皮部位呈強陽性表達(黑箭) DAB ×400;2F.PDR注藥組,VEGFR2蛋白在新生血管內皮細胞中呈弱陽性表達(黑箭)DAB ×400
圖2
PDR患眼纖維血管膜病理標本免疫組織化學染色像。2A.PDR未注藥組,Notch1蛋白在新生血管內皮細胞中呈強陽性表達,而在纖維細胞和細胞外基質中幾乎無表達(黑箭) DAB ×400;2B.PDR注藥組,Notch1蛋白在血管內皮細胞中呈弱陽性表達(黑箭) DAB ×400;2C.PDR未注藥組,Dll4蛋白僅在新生血管內皮細胞中呈陽性表達,DAB ×400;2D.PDR注藥組,Dll4蛋白在血管內皮細胞中呈弱陽性表達(黑箭)DAB ×400;2E.PDR未注藥組,VEGFR2蛋白在新生血管內皮部位呈強陽性表達(黑箭) DAB ×400;2F.PDR注藥組,VEGFR2蛋白在新生血管內皮細胞中呈弱陽性表達(黑箭)DAB ×400
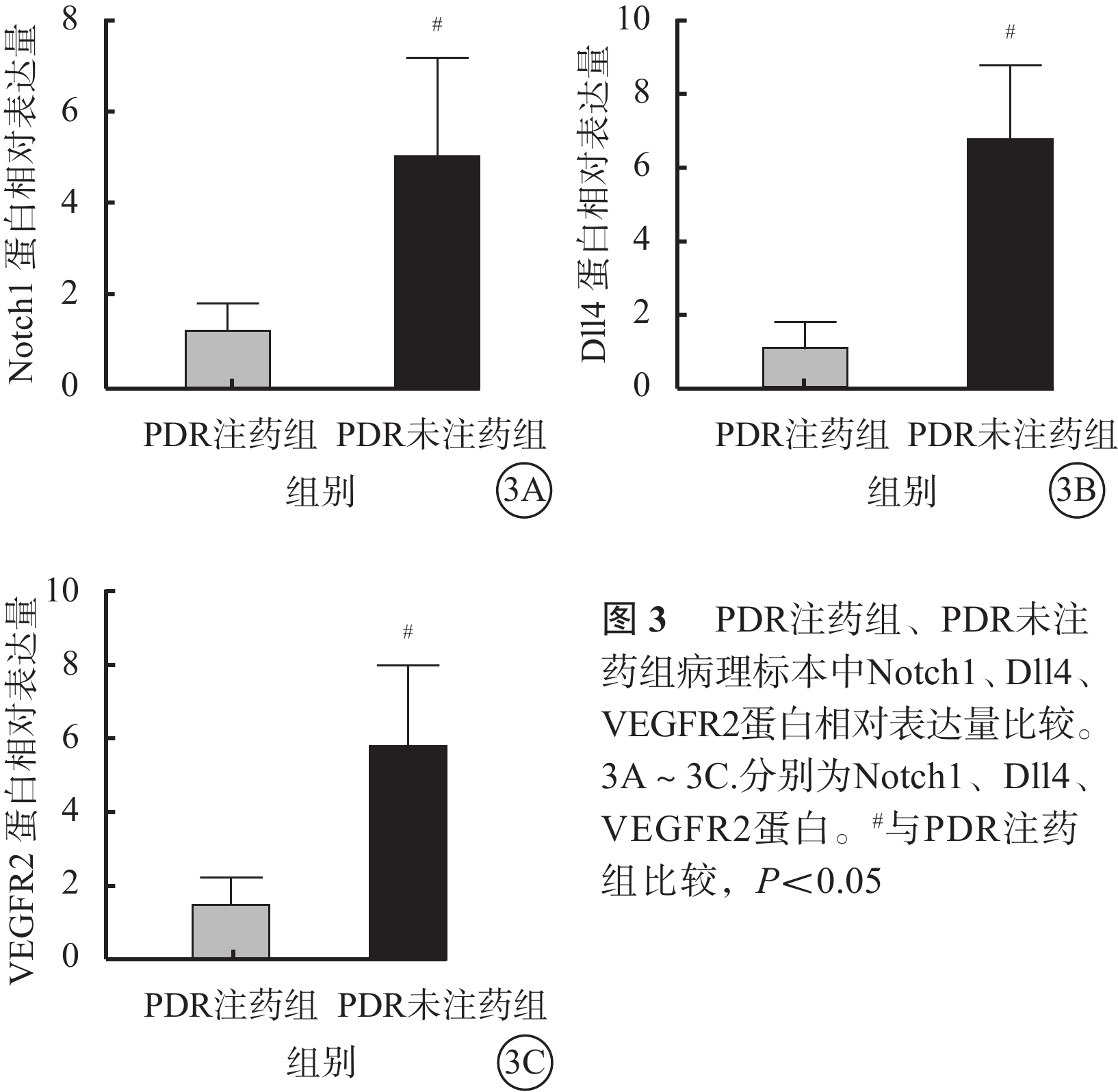
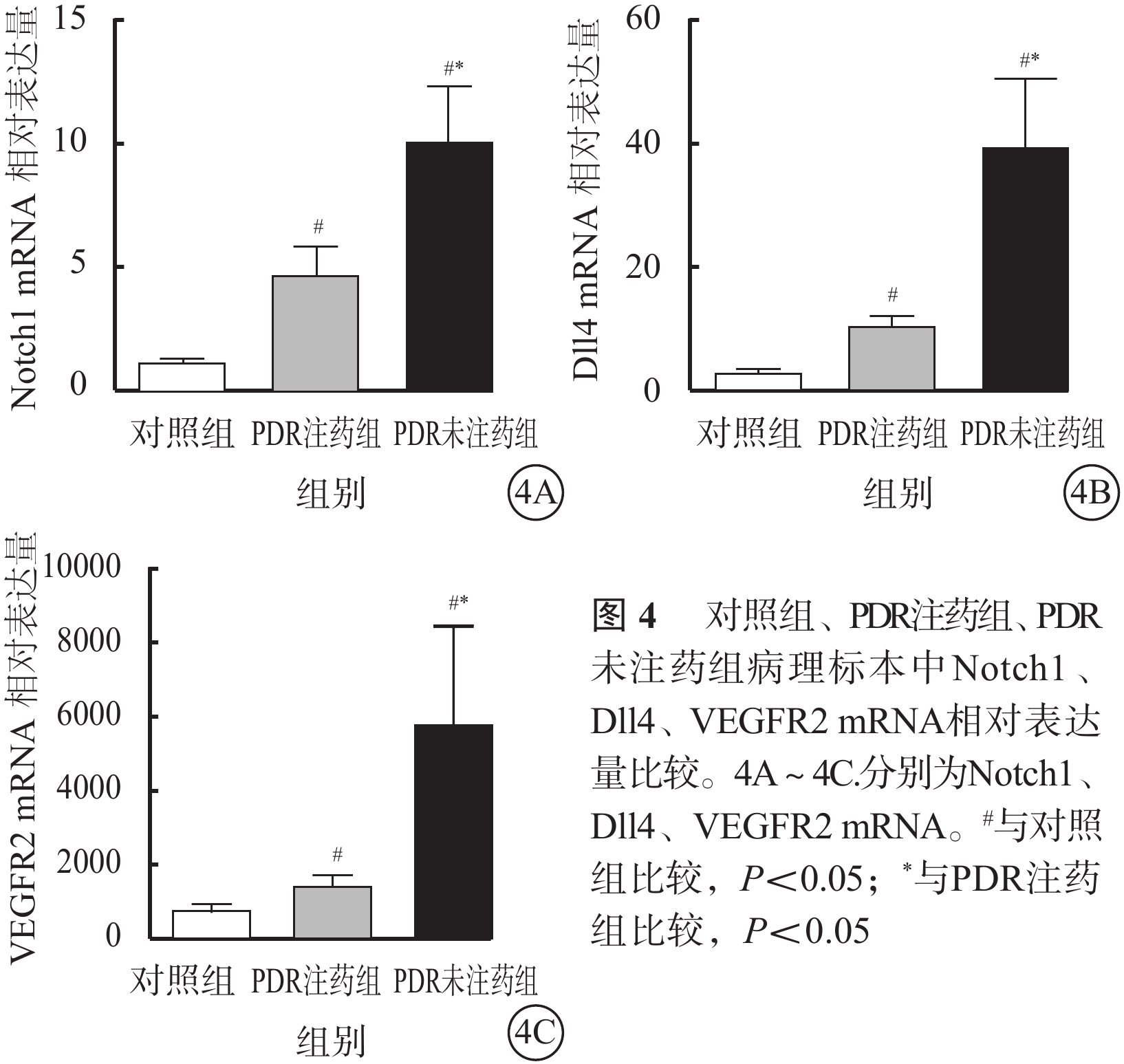
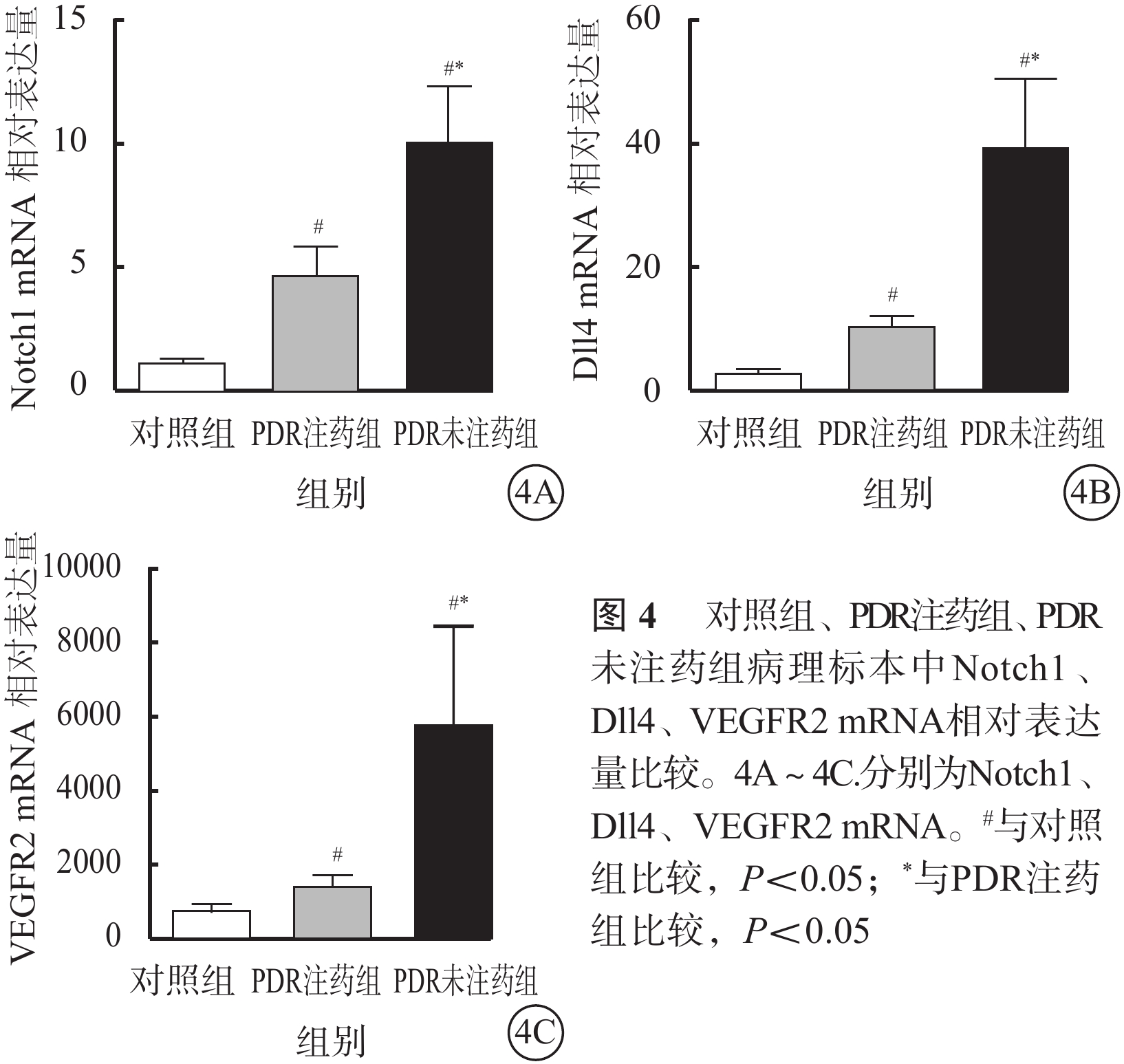
RT-PCR檢測結果顯示,PDR注藥組(H=12.50)、PDR未注藥組(H=12.50)、對照組(H=12.02)纖維血管膜、黃斑前膜病理標本中Notch1、Dll4、VEGFR2 mRNA相對表達量比較,差異均有統計學意義(P=0.002、0.002、0.002)(圖4)。其中,PDR未注藥組纖維血管膜病理標本中Notch1、Dll4、VEGFR2 mRNA相對表達量最高,對照組黃斑前膜病理標本中相對表達量最低。對照組與PDR注藥組、PDR注藥組與PDR未注藥組、對照組與PDR未注藥組病理標本中Notch1、Dll4、VEGFR2 mRNA相對表達量比較,差異均有統計學意義(P<0.05)。
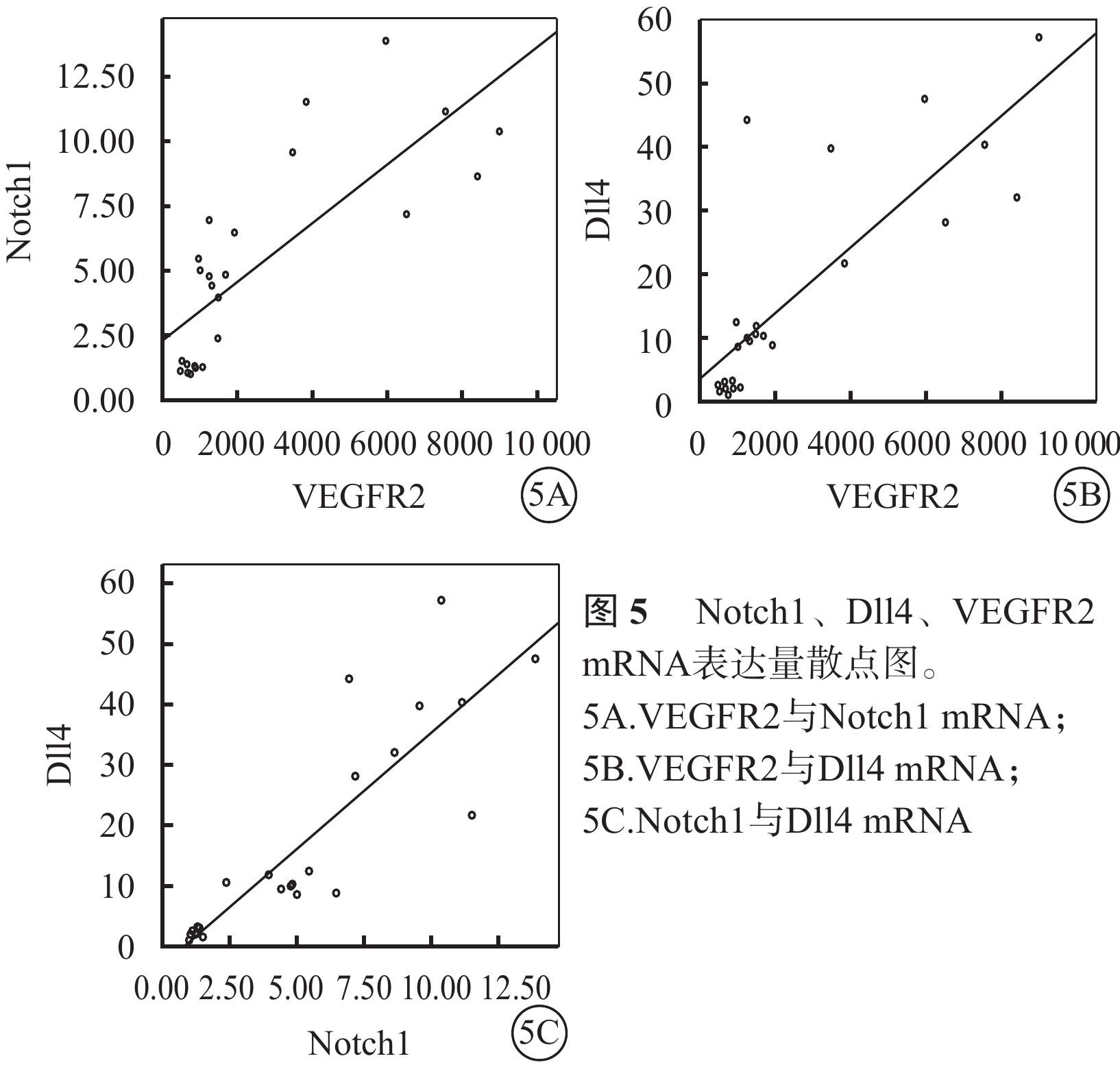
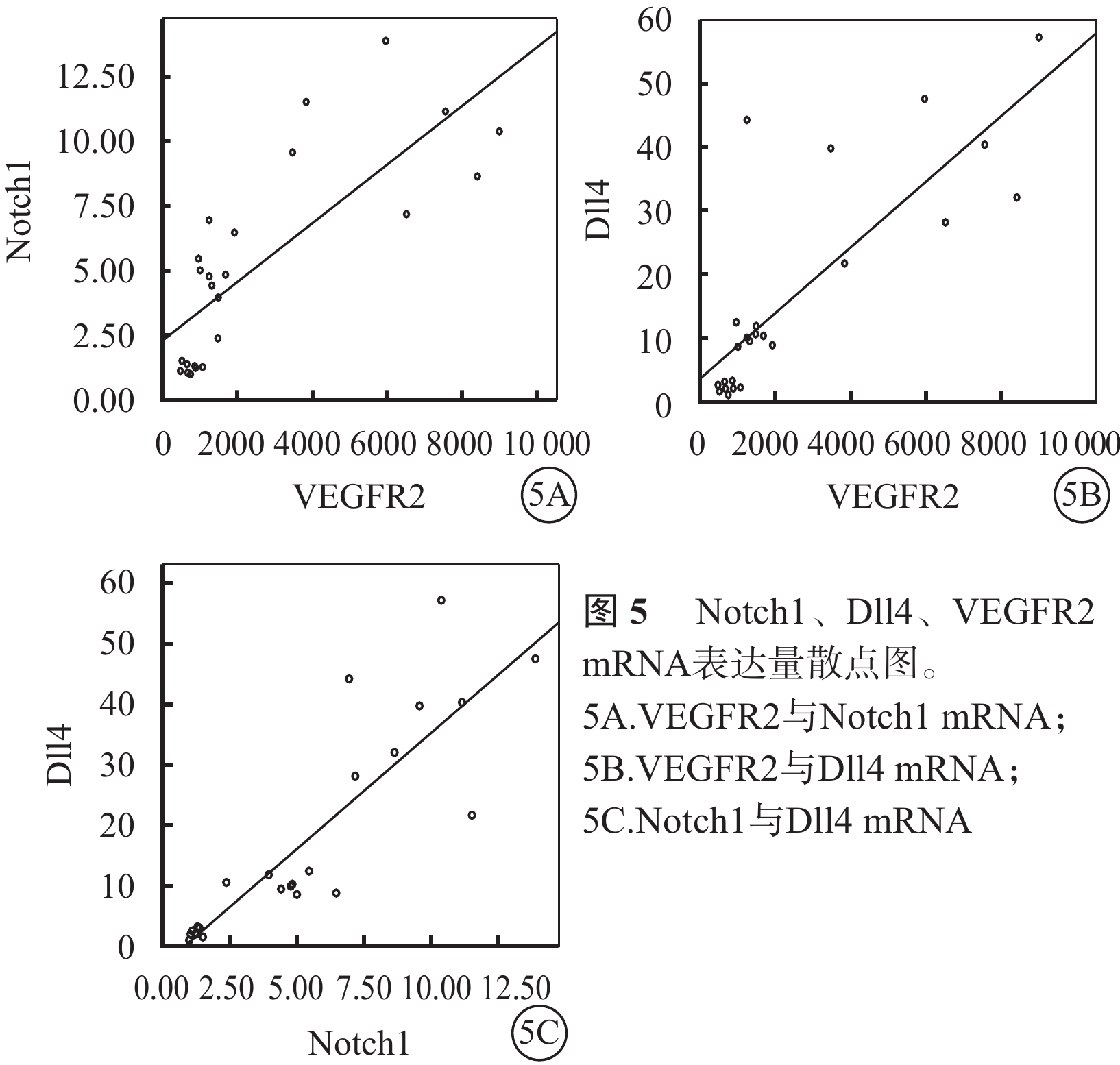
Spearman相關分析結果顯示,VEGFR2與Notch1(r=0.83)、VEGFR2與Dll4(r=0.81)、Notch1與Dll4(r=0.87) mRNA的表達呈顯著正相關(P<0.05)(圖5)。
3 討論
Notch信號通路是一條在脊椎動物和無脊椎動物中均高度保守的信號傳導通路。研究發現,新生血管內皮細胞中表達Notch信號通路中的Notch1、Notch4兩種受體及Jagged1、Dll1、Dll4三種配體[8]。其中,Dll4集中表達在胚胎及成年組織的血管內皮細胞中,在血管重構中發揮重要作用[9],Dll4/Notch基因可促進新生血管的成熟[10]。實驗研究發現,阻斷Notch信號通路可以減少新生血管的生成[11],而Wang等[12]在對卵巢癌患者組織的研究中證實,Notch1、Dll4的表達量顯著高于正常對照組,因此認為Dll4可用于檢測對卵巢癌新生血管的治療效果。眼內研究同樣表明,Dll4在發育中的小鼠視網膜尖端細胞上呈現高水平表達,而在莖細胞中表達不明顯[13]。
PDR以眼底新生血管形成為最主要標志,并形成大量纖維血管膜,可導致玻璃體積血及后期牽拉性視網膜脫離的發生,是以缺氧導致的視網膜前新生血管形成為最主要病理改變的眼底病變。在PDR中,由于視網膜微血管細胞損害,造成視網膜微血管閉塞、無灌注區形成,從而引起局部組織缺血、缺氧。而越來越多的研究表明,缺氧也被認為是Notch信號通路的重要始動環節[14,15];在PDR新生血管形成過程中,Dll4/Notch1信號通路作用于動脈內皮,發揮調控血管重構,降低血管出芽及側支數量,促進新生血管分化成熟的作用。因此,PDR注藥組、PDR未注藥組纖維血管膜中Notch1、Dll4的表達量顯著高于對照組。PDR注藥組纖維血管膜中Notch1、Dll4的表達量低于未注藥組,分析其原因可能是由VEGF信號通路對Dll4/Notch信號通路的調節所致。Dll4/Notch與VEGF兩個信號通路是通過反饋調節機制調節血管新生的。VEGF的高表達可誘導不同來源的多基因表達,其中就包括受體Notch1及配體Dll4基因的轉錄和表達[16]。Ridgway等[17]通過向玻璃體腔中注射VEGF蛋白,發現視網膜Dll4表達及血管出芽明顯增加。而在腫瘤內皮細胞中,當用VEGF Trap阻斷VEGF信號通路后,Dll4的表達水平顯著降低,說明Dll4在腫瘤內皮細胞中的表達依賴于VEGF[3]。本研究中,玻璃體腔注射抗VEGF藥物后,VEGF信號通路的抑制和阻斷減弱了對Notch信號通路的正向調節作用;另一方面,抗VEGF藥物的玻璃體腔注射抑制了活動性血管的生長,從而使部分視網膜新生血管的生長明顯衰退。因此,位于新生血管內皮細胞中的Notch1、Dll4亦相應減少。
VEGF是目前研究最多的促血管生成因子。通常認為,VEGF與VEGFR2之間的信號傳遞是VEGF信號通路中最主要的介導途徑。Smith等[18]在對DR患者的研究中發現,VEGFR2表達量的高低可間接反映眼底血管新生的程度強弱。因此在本研究中,我們通過測量VEGFR2的表達檢測VEGF信號通路的表達活性。結果發現PDR注藥組、PDR未注藥組患眼病理標本中VEGFR2表達量相對于對照組升高,考慮可能由于對照組黃斑前膜組織并未有新生血管的出芽和形成,因此亦未有VEGF信號通路的過度激活。PDR注藥組纖維血管膜組織中VEGFR2、Notch1、Dll4 mRNA及其蛋白相對表達量較PDR未注藥組均降低,且Notch1、Dll4的表達與VEGFR2的表達均呈正相關,免疫組織化學染色結果顯示,VEGFR2的表達部位與Dll4、Notch1的表達部位大體一致,均主要表達于新生血管內皮組織,說明Notch1、Dll4的表達受VEGF信號通路的影響,共同調節PDR新生血管的形成。此外,受檢眼病理標本中新生血管內皮細胞數目PDR未注藥組也大于PDR注藥組,差異有統計學意義,分析可能由于PDR未注藥組VEGF濃度高于PDR注藥組。這與前述Notch信號通路可促進病理性新生血管的發生和成熟的結論吻合。
結合目前國內外對Notch信號通路已有的研究,本研究結果提示,Notch信號通路可能在PDR的新生血管形成和發展中發揮一定作用,并且該作用受VEGF信號通路的誘導和調控。DR是一個多因素共同作用的非常復雜的過程,而探索Notch信號通路在DR發病過程中的表達,有助于進一步研究DR的發病機制,為臨床防治DR提供新的思路和靶點。
讀者?作者?編者 關于作者署名以及單位著錄的要求 論文作者署名以及單位著錄是一嚴肅慎重的事情。盡管本刊根據中華醫學會雜志應以及其他相關規定的要求,對其反復強調和說明,但在論文投稿、修稿和校對過程中,仍有不少與作者署名以及單位著錄相關的不當事情發生。為此,再將本刊對論文作者署名以及單位著錄的要求重申如下,請作者務必遵照執行。對于不符合規定及要求的文稿,不能進入本刊稿件處理流程;本刊有權拒絕并提請作者進行相應的說明或修改。 1 作者署名 參與選題設計或參與資料分析解釋,起草修改論文中關鍵性理論或其他主要內容,能對編輯部修稿意見進行審核修改并在學術界進行答辯并且最終同意該文發表者方能作為文稿的作者進行署名。僅參與獲得資金或收集資料者不能列為作者;僅對科研小組進行一般管理者也不宜列為作者。對研究有貢獻但不宜作為作者署名的其他人員可在文后志謝處列出。 每篇文稿作者中需要確定1名能對該論文全面負責的通信作者。通信作者應在投稿時確定,如在來稿中未特殊標明,則視第一作者為通信作者。第一作者與通信作者不是同一人時,應在論文首頁腳注通信作者姓名以及電子郵件。需要特別提請注意的是,本刊目前沒有采納并列第一作者及并列通信作者的作者署名方式。作者中如有外籍作者,應附外籍作者本人親筆簽名同意署名文稿在本刊發表的函件。集體署名的論文于文題下列署名集體,于文末列整理者姓名并于集體署名下列出通信作者姓名、單位和郵政編碼。集體署名的文章必須將對該文負責的關鍵人物列為通信作者。文稿第一作者在投稿后工作單位有變動,應同時列出其原單位及現在單位。 研究生、進修醫生在學習和進修期間利用在讀學校或進修醫院的條件、設備、病例資料完成的論文,在征得其單位同意后可以以作者身份參與署名,但體現知識產權歸屬的作者單位應著錄為該學校或醫院。不能將研究生畢業后工作單位或進修醫生歸屬單位著錄為作者單位;也不能把現在所在單位的其他無關人員列為作者參與署名。 作者署名時,作者姓名在文題下按序排列,排序應在投稿時確定。在投稿后的稿件處理編排過程中不應再做更改。必須更改順序或增刪者,應附所有作者親筆簽名同意以及單位簽署公章的更改說明。 2 單位著錄 提供研究條件并實際完成研究工作的單位才能作為文稿知識產權的享有者著錄為作者單位。不同單位共同撰寫的文稿,各個單位的名稱均須分別列出,由第一作者所在單位開具論文推薦信。文稿為集體作者,應由第一整理者(即第一執筆者)所在單位開具論文推薦信。研究生、進修醫生在學習和進修期間,利用學習和進修單位的條件和資料完成的工作,第一作者單位應署提供條件和資料的單位,但可以注明第一作者現在所在單位。聯合培養的研究生其稿件中單位署名及排名順序可按所在學校、單位要求著錄,但應在投稿時予以說明。 單位名稱應按文稿知識產權單位組織機構代碼中的規范名稱著錄;與論文推薦信中單位公章中的標準名稱保持一致。同一單位具有不同公章的多個名稱者,不同名稱的著錄以公章中的名稱為準;不同名稱之間的排序應在投稿時確定并在論文推薦信中簽署相應的公章。必要時應對單位署名和排序的特殊要求另予說明。已歸屬于綜合大學的單位,應先列出大學名稱,之后列出醫學院名稱或醫院名稱、科室名稱。科室有多種名稱者,應按所在單位的要求使用規范的名稱。單位的英文名稱應根據所在單位統一的英文名稱書寫。 本刊編輯部
Notch信號通路在血管生成過程中發揮重要作用[1]。除了維持視網膜正常生理微環境,其缺失將導致視網膜血管和細胞發育不良及視網膜環境穩態失調[2],并且參與缺氧條件下視網膜新生血管的發生[3]。作為Notch信號通路的配體之一,Delta樣配體(Dll)4已被證實可特異性表達于生理及病理新生血管的發生中[4]。我們前期動物模型研究發現,Notch受體蛋白1(Notch1)、Dll4的表達量在早期糖尿病視網膜病變(DR)發展過程中逐漸升高,血管內皮生長因子(VEGF)受體2(VEGFR2)表達量逐漸增加,且VEGFR2的表達與Notch1、Dll4呈正相關[5]。但Notch信號通路是否參與人類增生型DR(PDR)新生血管的發生尚不清楚。因此,我們通過檢測PDR患眼纖維血管膜病理標本中Notch1、Dll4、VEGFR2的表達,進一步探討Notch1、Dll4在PDR新生血管形成中的作用及與VEGF信號通路的關系。現將結果報道如下。
1 對象和方法
2014年7月至2015年7月在青島大學附屬醫院眼科行睫狀體平坦部23G標準三通道玻璃體切割手術的PDR患者57例60只眼納入研究。本研究獲得青島大學附屬醫院倫理審查委員會審查批準;納入研究對象或家屬均簽署知情同意書。患者中男性32例34只眼,女性25例26只眼。年齡36~73歲,平均年齡(54.93±6.42)歲。糖尿病病程8~20年,平均糖尿病病程(15.72±3.51)年。所有患者均符合2型糖尿病診斷標準及PDR診斷標準[6,7]。排除肺部疾病、嚴重肝腎功能不全、心腦血管病變、惡性腫瘤及自身免疫性疾病者。
根據手術前是否行抗VEGF藥物玻璃體腔注射將患眼分為PDR未注藥組和PDR注藥組,分別為30例32只眼、27例28只眼。PDR未注藥組30例32只眼中,男性16例18只眼,女性14例14只眼;年齡36~73歲,平均年齡55歲;糖尿病病程8~20年,平均糖尿病病程16年。PDR注藥組27例28只眼中,男性16例16只眼,女性11例12只眼;年齡40~68歲,平均年齡54歲;糖尿病病程8~18年,平均糖尿病病程15年。玻璃體切割手術前2~7 d行雷珠單抗玻璃體腔注射治療。選取特發性黃斑前膜患者18例18只眼作為對照組。其中,男性5例5只眼,女性13例13只眼;年齡43~79歲,平均年齡61歲。均無糖尿病及糖尿病家族史,眼部及全身感染、急性炎癥反應及缺血缺氧性疾病史以及全身系統性疾病。3組患者年齡(F=1.14)、性別構成比(F=0.97)比較,差異均無統計學意義(P>0.05)。
玻璃體切割手術中應用視網膜剪及視網膜鑷獲得PDR纖維血管膜病理標本及黃斑前膜病理標本。隨機數字表法隨機分配標本。30只眼病理標本立即用4%多聚甲醛溶液固定,常規酒精脫水,二甲苯透明,浸蠟包埋,3 μm連續切片并編號,行光學顯微鏡觀察及蛋白表達檢測;48只眼病理標本則移入已消毒的離心管中,–80 ℃超低溫冰箱中凍存,用于基因檢測。行光學顯微鏡觀察及蛋白表達檢測的30只眼病理標本中,PDR未注藥組、PDR注藥組、對照組分別為10、12、8只眼;行基因檢測的48只眼病理標本中,PDR未注藥組、PDR注藥組、對照組分別為22、16、10只眼。
所有石蠟標本截面最大處連續切片5張進行蘇木精-伊紅(HE)染色。高倍光學顯微鏡下觀察。每張切片隨機選取5個視野采用盲法計數突破內界膜的血管內皮細胞,各切片內皮細胞計數為5個視野中細胞計數的平均值,最后計算各組病理標本每張切片的血管內皮細胞計數的平均值。
免疫組織化學方法檢測各組病理標本中Notch1、Dll4、VEGFR2蛋白表達。每個病理標本選取截面最大處連續切片15張,各選取5張按照免疫組織化學試劑盒說明要求進行Notch1、Dll4及VEGFR2蛋白的免疫組織化學染色,二氨基苯聯胺(DAB)顯色3 min,磷酸鹽緩沖液代替一抗作為陰性對照。結果以細胞漿內出現黃色、棕黃色或棕褐色顆粒為弱陽性、陽性、強陽性。采用Image pro plus6.0軟件計算每張切片相應蛋白免疫組織化學染色陽性部分的平均吸光度[A,舊稱光密度(OD)]值,最后統計各組病理標本中各種蛋白的平均值,代表相應蛋白的表達量。
熒光定量聚合酶鏈反應(RT-PCR)檢測各組病理標本中Notch1、Dll4、VEGFR2 mRNA的表達。50~100 mg纖維血管膜或黃斑前膜加入1 ml RNAiso plus進行裂解;提取總RNA,紫外分光光度計檢測其濃度和純度,A260/A280在1.7~2.1的樣本用于試驗。逆轉錄成cDNA,以磷酸甘油醛脫氫酶(GAPDH)作為內參基因,采用雙鏈嵌合熒光染色法擴增相應基因。引物序列:Notch1:上游引物5′-GTCAACGCCGTAGAT-GACC-3′,下游引物5′-TTGTTAGCCCCGTTCTTCAG-3′,長度101堿基對(bp);Dll4:上游引物5′-CCC-TGGCAATGTACTTGTGAT-3′,下游引物5′-TGGT-GGGTGCAGTAGTTGAG-3,長度74 bp;VEGFR2:上游引物5′-AGTGATCGGAAATGACACTGGA-3′,下游引物5′-GCACAAAGTGACACGTTGAGAT-3′,長度213 bp;GAPDH:上游引物5′-CACGATGGAGGGG-CCGGACTCATC-3′,下游引物5′-TAAAGACCTCTAT-GCCAACACAGT -3′,長度241 bp。退火溫度60 ℃,40個循環后形成擴增曲線,記錄循環閾值(Ct值),結果采用2–ΔΔCt公式進行分析。
采用SPSS19.0統計學軟件進行統計學分析處理。數據經Kolmogoror-Smimor檢驗證實成正態分布后用均數±標準差( )表示。多組間相關因子表達比較采用Kruskal-Wallis檢驗。兩變量間關系采用Spearman相關性分析。α=0.05。P<0.05為差異有統計學意義。
2 結果
光學顯微鏡觀察發現,PDR纖維血管膜病理標本均可見新生血管生成,但新生血管數量和形態不同。PDR注藥組12只眼纖維血管膜病理標本中可見血管管腔較狹窄,部分管腔出現閉合。PDR未注藥組10只眼纖維血管膜病理標本可見血管管腔擴張,形態較完整。對照組8只眼黃斑前膜病理標本中可見各種細胞散布其中,未見血管結構;未見突破內界膜的血管內皮細胞(圖1)。PDR注藥組、PDR未注藥組纖維血管膜病理標本中突破內界膜的血管內皮細胞數量分別為(17.17±2.48)、(41.50±5.57)個;兩組纖維血管膜病理標本突破內界膜的血管內皮細胞數量比較,差異有統計學意義(t=9.58,P<0.05)。
免疫組織化學染色結果顯示,PDR 22只眼纖維血管膜病理標本中Notch1、Dll4、VEGFR2表達均呈陽性,且主要表達在新生血管內皮組織,血管外部位表達較少(圖2)。PDR注藥組、PDR未注藥組纖維血管膜病理標本中Notch1、Dll4、VEGFR2蛋白相對表達量比較,差異均有統計學意義(t=3.45、6.01、4.08,P=0.030、0.008、0.023)(圖3)。
圖2
PDR患眼纖維血管膜病理標本免疫組織化學染色像。2A.PDR未注藥組,Notch1蛋白在新生血管內皮細胞中呈強陽性表達,而在纖維細胞和細胞外基質中幾乎無表達(黑箭) DAB ×400;2B.PDR注藥組,Notch1蛋白在血管內皮細胞中呈弱陽性表達(黑箭) DAB ×400;2C.PDR未注藥組,Dll4蛋白僅在新生血管內皮細胞中呈陽性表達,DAB ×400;2D.PDR注藥組,Dll4蛋白在血管內皮細胞中呈弱陽性表達(黑箭)DAB ×400;2E.PDR未注藥組,VEGFR2蛋白在新生血管內皮部位呈強陽性表達(黑箭) DAB ×400;2F.PDR注藥組,VEGFR2蛋白在新生血管內皮細胞中呈弱陽性表達(黑箭)DAB ×400
RT-PCR檢測結果顯示,PDR注藥組(H=12.50)、PDR未注藥組(H=12.50)、對照組(H=12.02)纖維血管膜、黃斑前膜病理標本中Notch1、Dll4、VEGFR2 mRNA相對表達量比較,差異均有統計學意義(P=0.002、0.002、0.002)(圖4)。其中,PDR未注藥組纖維血管膜病理標本中Notch1、Dll4、VEGFR2 mRNA相對表達量最高,對照組黃斑前膜病理標本中相對表達量最低。對照組與PDR注藥組、PDR注藥組與PDR未注藥組、對照組與PDR未注藥組病理標本中Notch1、Dll4、VEGFR2 mRNA相對表達量比較,差異均有統計學意義(P<0.05)。
Spearman相關分析結果顯示,VEGFR2與Notch1(r=0.83)、VEGFR2與Dll4(r=0.81)、Notch1與Dll4(r=0.87) mRNA的表達呈顯著正相關(P<0.05)(圖5)。
3 討論
Notch信號通路是一條在脊椎動物和無脊椎動物中均高度保守的信號傳導通路。研究發現,新生血管內皮細胞中表達Notch信號通路中的Notch1、Notch4兩種受體及Jagged1、Dll1、Dll4三種配體[8]。其中,Dll4集中表達在胚胎及成年組織的血管內皮細胞中,在血管重構中發揮重要作用[9],Dll4/Notch基因可促進新生血管的成熟[10]。實驗研究發現,阻斷Notch信號通路可以減少新生血管的生成[11],而Wang等[12]在對卵巢癌患者組織的研究中證實,Notch1、Dll4的表達量顯著高于正常對照組,因此認為Dll4可用于檢測對卵巢癌新生血管的治療效果。眼內研究同樣表明,Dll4在發育中的小鼠視網膜尖端細胞上呈現高水平表達,而在莖細胞中表達不明顯[13]。
PDR以眼底新生血管形成為最主要標志,并形成大量纖維血管膜,可導致玻璃體積血及后期牽拉性視網膜脫離的發生,是以缺氧導致的視網膜前新生血管形成為最主要病理改變的眼底病變。在PDR中,由于視網膜微血管細胞損害,造成視網膜微血管閉塞、無灌注區形成,從而引起局部組織缺血、缺氧。而越來越多的研究表明,缺氧也被認為是Notch信號通路的重要始動環節[14,15];在PDR新生血管形成過程中,Dll4/Notch1信號通路作用于動脈內皮,發揮調控血管重構,降低血管出芽及側支數量,促進新生血管分化成熟的作用。因此,PDR注藥組、PDR未注藥組纖維血管膜中Notch1、Dll4的表達量顯著高于對照組。PDR注藥組纖維血管膜中Notch1、Dll4的表達量低于未注藥組,分析其原因可能是由VEGF信號通路對Dll4/Notch信號通路的調節所致。Dll4/Notch與VEGF兩個信號通路是通過反饋調節機制調節血管新生的。VEGF的高表達可誘導不同來源的多基因表達,其中就包括受體Notch1及配體Dll4基因的轉錄和表達[16]。Ridgway等[17]通過向玻璃體腔中注射VEGF蛋白,發現視網膜Dll4表達及血管出芽明顯增加。而在腫瘤內皮細胞中,當用VEGF Trap阻斷VEGF信號通路后,Dll4的表達水平顯著降低,說明Dll4在腫瘤內皮細胞中的表達依賴于VEGF[3]。本研究中,玻璃體腔注射抗VEGF藥物后,VEGF信號通路的抑制和阻斷減弱了對Notch信號通路的正向調節作用;另一方面,抗VEGF藥物的玻璃體腔注射抑制了活動性血管的生長,從而使部分視網膜新生血管的生長明顯衰退。因此,位于新生血管內皮細胞中的Notch1、Dll4亦相應減少。
VEGF是目前研究最多的促血管生成因子。通常認為,VEGF與VEGFR2之間的信號傳遞是VEGF信號通路中最主要的介導途徑。Smith等[18]在對DR患者的研究中發現,VEGFR2表達量的高低可間接反映眼底血管新生的程度強弱。因此在本研究中,我們通過測量VEGFR2的表達檢測VEGF信號通路的表達活性。結果發現PDR注藥組、PDR未注藥組患眼病理標本中VEGFR2表達量相對于對照組升高,考慮可能由于對照組黃斑前膜組織并未有新生血管的出芽和形成,因此亦未有VEGF信號通路的過度激活。PDR注藥組纖維血管膜組織中VEGFR2、Notch1、Dll4 mRNA及其蛋白相對表達量較PDR未注藥組均降低,且Notch1、Dll4的表達與VEGFR2的表達均呈正相關,免疫組織化學染色結果顯示,VEGFR2的表達部位與Dll4、Notch1的表達部位大體一致,均主要表達于新生血管內皮組織,說明Notch1、Dll4的表達受VEGF信號通路的影響,共同調節PDR新生血管的形成。此外,受檢眼病理標本中新生血管內皮細胞數目PDR未注藥組也大于PDR注藥組,差異有統計學意義,分析可能由于PDR未注藥組VEGF濃度高于PDR注藥組。這與前述Notch信號通路可促進病理性新生血管的發生和成熟的結論吻合。
結合目前國內外對Notch信號通路已有的研究,本研究結果提示,Notch信號通路可能在PDR的新生血管形成和發展中發揮一定作用,并且該作用受VEGF信號通路的誘導和調控。DR是一個多因素共同作用的非常復雜的過程,而探索Notch信號通路在DR發病過程中的表達,有助于進一步研究DR的發病機制,為臨床防治DR提供新的思路和靶點。
讀者?作者?編者 關于作者署名以及單位著錄的要求 論文作者署名以及單位著錄是一嚴肅慎重的事情。盡管本刊根據中華醫學會雜志應以及其他相關規定的要求,對其反復強調和說明,但在論文投稿、修稿和校對過程中,仍有不少與作者署名以及單位著錄相關的不當事情發生。為此,再將本刊對論文作者署名以及單位著錄的要求重申如下,請作者務必遵照執行。對于不符合規定及要求的文稿,不能進入本刊稿件處理流程;本刊有權拒絕并提請作者進行相應的說明或修改。 1 作者署名 參與選題設計或參與資料分析解釋,起草修改論文中關鍵性理論或其他主要內容,能對編輯部修稿意見進行審核修改并在學術界進行答辯并且最終同意該文發表者方能作為文稿的作者進行署名。僅參與獲得資金或收集資料者不能列為作者;僅對科研小組進行一般管理者也不宜列為作者。對研究有貢獻但不宜作為作者署名的其他人員可在文后志謝處列出。 每篇文稿作者中需要確定1名能對該論文全面負責的通信作者。通信作者應在投稿時確定,如在來稿中未特殊標明,則視第一作者為通信作者。第一作者與通信作者不是同一人時,應在論文首頁腳注通信作者姓名以及電子郵件。需要特別提請注意的是,本刊目前沒有采納并列第一作者及并列通信作者的作者署名方式。作者中如有外籍作者,應附外籍作者本人親筆簽名同意署名文稿在本刊發表的函件。集體署名的論文于文題下列署名集體,于文末列整理者姓名并于集體署名下列出通信作者姓名、單位和郵政編碼。集體署名的文章必須將對該文負責的關鍵人物列為通信作者。文稿第一作者在投稿后工作單位有變動,應同時列出其原單位及現在單位。 研究生、進修醫生在學習和進修期間利用在讀學校或進修醫院的條件、設備、病例資料完成的論文,在征得其單位同意后可以以作者身份參與署名,但體現知識產權歸屬的作者單位應著錄為該學校或醫院。不能將研究生畢業后工作單位或進修醫生歸屬單位著錄為作者單位;也不能把現在所在單位的其他無關人員列為作者參與署名。 作者署名時,作者姓名在文題下按序排列,排序應在投稿時確定。在投稿后的稿件處理編排過程中不應再做更改。必須更改順序或增刪者,應附所有作者親筆簽名同意以及單位簽署公章的更改說明。 2 單位著錄 提供研究條件并實際完成研究工作的單位才能作為文稿知識產權的享有者著錄為作者單位。不同單位共同撰寫的文稿,各個單位的名稱均須分別列出,由第一作者所在單位開具論文推薦信。文稿為集體作者,應由第一整理者(即第一執筆者)所在單位開具論文推薦信。研究生、進修醫生在學習和進修期間,利用學習和進修單位的條件和資料完成的工作,第一作者單位應署提供條件和資料的單位,但可以注明第一作者現在所在單位。聯合培養的研究生其稿件中單位署名及排名順序可按所在學校、單位要求著錄,但應在投稿時予以說明。 單位名稱應按文稿知識產權單位組織機構代碼中的規范名稱著錄;與論文推薦信中單位公章中的標準名稱保持一致。同一單位具有不同公章的多個名稱者,不同名稱的著錄以公章中的名稱為準;不同名稱之間的排序應在投稿時確定并在論文推薦信中簽署相應的公章。必要時應對單位署名和排序的特殊要求另予說明。已歸屬于綜合大學的單位,應先列出大學名稱,之后列出醫學院名稱或醫院名稱、科室名稱。科室有多種名稱者,應按所在單位的要求使用規范的名稱。單位的英文名稱應根據所在單位統一的英文名稱書寫。 本刊編輯部