引用本文: 侯亞南, 雷寧玉, 宋斌, 王麗君. 音猬因子信號通路在缺氧誘導的人視網膜色素上皮細胞中血管內皮生長因子表達中的作用. 中華眼底病雜志, 2016, 32(1): 62-65. doi: 10.3760/cma.j.issn.1005-1015.2016.01.015 復制
血管內皮生長因子(VEGF)為重要的促新生血管生成因子, 參與缺血缺氧誘導的視網膜、脈絡膜新生血管的形成[1-3]。視網膜周細胞、微血管內皮細胞、Müller細胞和視網膜色素上皮(RPE)細胞皆可產生VEGF, 其中以RPE細胞表達VEGF能力最為顯著[4]。Hedgehog (Hh)信號通路在調控脊椎動物細胞增生和血管發生、生成中起到重要作用[5]。脊椎動物Hh基因包含音猬因子(Shh)、沙漠刺猬因子(Dhh)、印度刺猬因子(Ihh),其中Shh信號與血管發育密切相關[6]。研究結果表明,眾多腫瘤的發生與Shh信號通路相關[7-12],并且已有臨床數據證實腫瘤組織中Shh信號通路與VEGF的表達存在相關性,但兩者具體作用機制尚不明確[13]。近期研究結果顯示, 缺氧可上調人RPE(hRPE)細胞中VEGF的表達[14-16],但其信號轉導機制尚不完全清楚。為此,本研究初步探討Shh信號轉導通路在缺氧誘導hRPE細胞VEGF表達中的作用。現將結果報道如下。
1 材料和方法
hRPE-19(ARPE-19)細胞株(中山大學動物實驗中心);Dulbecco改良Eagle(DMEM)培養基、青鏈霉素、胰蛋白酶[中科邁晨(北京)科技有限公司];胎牛血清(杭州四季青生物工程材料有限公司);cyclopamine、CoCl2(美國Sigma公司);Trizol試劑,逆轉錄(RT)試劑盒、熒光定量聚合酶鏈反應(RT-PCR)試劑盒(大連寶生物工程技術服務有限公司);人VEGF酶鏈免疫吸附測定試驗(ELISA)試劑盒(上海依科賽生物技術公司)。
ARPE-19細胞株接種于含10%胎牛血清、100×103U/ml青霉素、100 mg/ml鏈霉素的DMEM培養液中,置于37℃,5%CO2的培養箱中培養,每3~4天換液傳代,取對數生長期3~6代細胞用于實驗。以細胞密度1×104、10×105、5×105個/孔分別接種于96、6、24孔板中,培養24 h后棄上清液,每孔加入無血清DMEM培養液饑餓24 h,將細胞分為正常對照組(Cont組)、缺氧組。缺氧組加入100μmol/L CoCl2后再分為缺氧對照組(CoCl2組)、cyclopamine處理組(CYA組)、二甲基亞砜對照組(DMSO組)。僅CYA組于造成缺氧前1 h加入20μmol/L的cyclopamine進行預處理[17, 18];DMSO組加入1‰DMSO。各組細胞培養4、8、12、24 h。
RT-PCR檢測各組細胞中Shh、VEGF mRNA的表達。取培養4、8、12、24 h的6孔板,分別提取各組細胞總RNA,逆轉錄成cDNA后進行擴增,紫外分光光度計測定RNA濃度為3.5~5.0 ng/μl,吸光度[A,舊稱光密度(OD)]260/A280為1.8~2.0。按照RT試劑盒說明書操作,反應體系為10μl。Shh、VEGF引物序列由大連寶生物工程技術服務有限公司合成。Shh擴增片段長度94堿基對(bp),上游引物:5′-TGT CTGCTGCTAGTCCTCGTCTC-3′,下游引物:5′-TT TTGGGGTGCCTCCTCTT-3′;VEGF擴增片段長度116 bp,上游引物:5′-CTGGAGTGTGTGCCCACTG A-3′,下游引物:5′-CATTCACATTTGTTGTGCTG TAGGA-3′;β-肌動蛋白(β-actin)擴增片段長度186 bp,上游引物:5′-TGGCACCCAGCACAATGAA-3′,下游引物:5′-CTAAGTCATAGTCCGCCTAGAAGCA-3′。擴增條件:94℃預變性30 s,95℃變性5 s,65℃退火30 s,39個循環;65℃延伸10 s。按照2-ΔΔCt計算各組VEGF、Shh mRNA相對表達量。實驗重復3次,取平均值。
ELISA檢測細胞培養上清液中VEGF蛋白含量。培養4、8、12、24 h的24孔板ARPE-19細胞分為Cont組、CoCl2組、CYA組、DMSO組,每組設3個副孔。取細胞培養上清液, 離心半徑4.5 cm,6000 r/min離心5 min。采用雙抗體夾心ELISA法,按照人VEGF ELISA試劑盒說明書進行操作。酶標儀450 nm波長讀取A值, 待測樣本的A值在標準曲線上查出該樣本的VEGF水平對應值。實驗重復3次,取平均值。
應用SPSS 13.0統計軟件行統計學分析處理。數據以均數±標準差(
2 結果
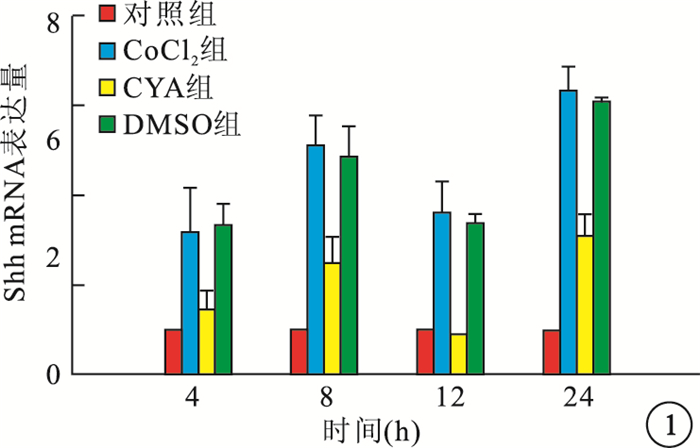
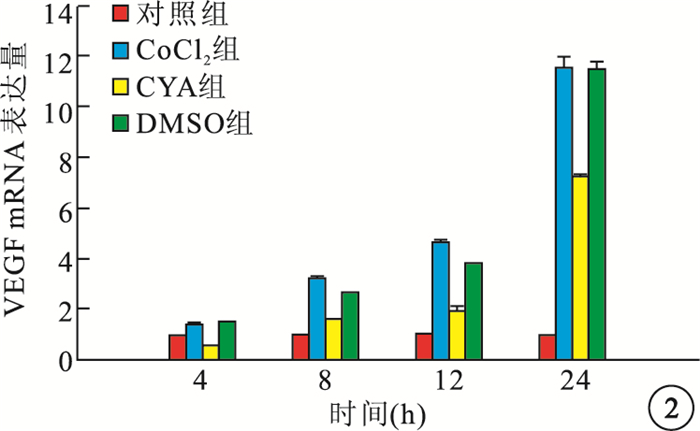
RT-PCR檢測結果顯示,缺氧組細胞內Shh、VEGF mRNA表達隨時間延長而逐漸增高,與Cont組細胞內Shh、VEGF mRNA表達比較,差異均有統計學意義(F=45.260、264.938,P=0.001、0.001)。CYA組細胞內Shh、VEGF mRNA表達較CoCl2組降低,差異均有統計學意義(P<0.01);DMSO組細胞內Shh、VEGF mRNA表達與CoCl2組比較,差異均無統計學意義(P>0.05)(圖 1, 2)。
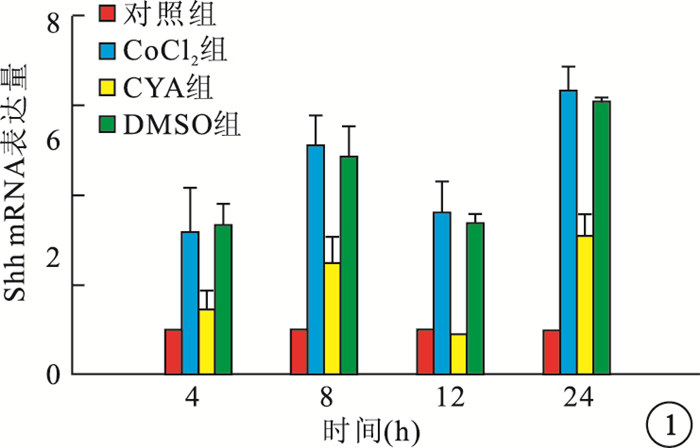 圖1
各組細胞內Shh mRNA表達比較
圖1
各組細胞內Shh mRNA表達比較
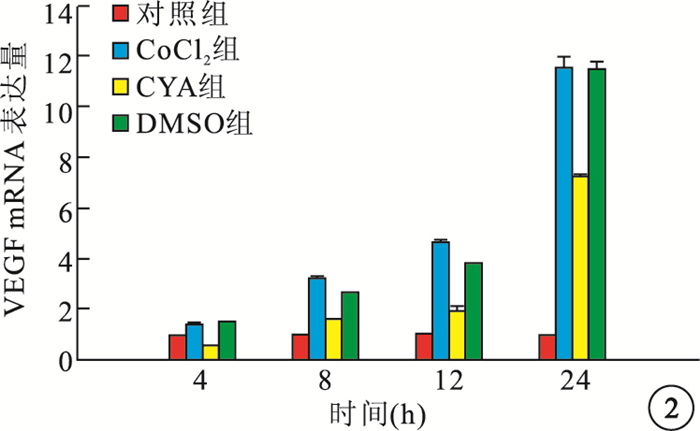 圖2
各組細胞內VEGF mRNA表達比較
圖2
各組細胞內VEGF mRNA表達比較
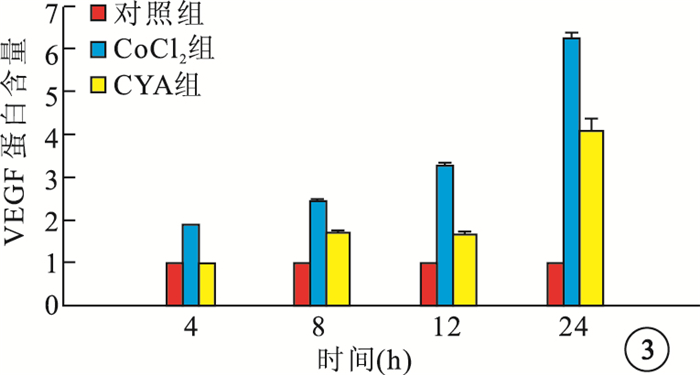
ELISA檢測結果顯示,CoCl2組細胞培養上清液中VEGF蛋白含量隨時間延長而顯著增加,與Cont組細胞培養上清液中VEGF蛋白含量比較,差異均有統計學意義(F=3 156.676,P=0.001);CYA組細胞培養上清液中VEGF蛋白含量較CoCl2組明顯降低, 差異均有統計學意義(P<0.01)(圖 3)。
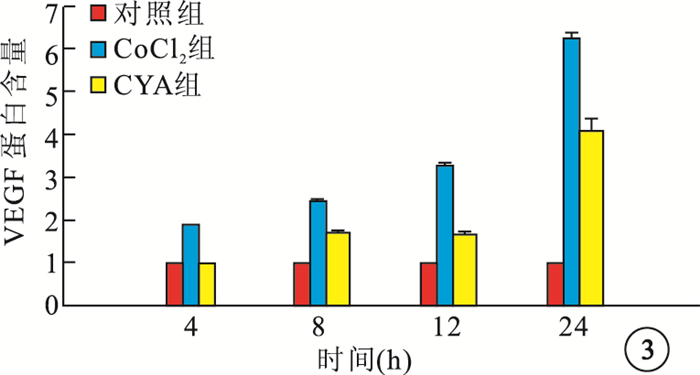 圖3
各組細胞培養上清液中VEGF蛋白含量比較
圖3
各組細胞培養上清液中VEGF蛋白含量比較
3 討論
Hh信號在血管發生和生成中起重要作用[19]。脊椎動物的Hh基因包含Shh、Dhh和Ihh[6],其中Shh信號與血管發育密切相關且對多種器官的形態發生分化不可或缺[19]。Shh被認為是一種脊椎動物體內間接的血管生成因子,可以直接作用于內皮細胞或刺激血管支持細胞產生上調VEGF和血管生成素[20]。既往研究發現,在生理條件下,外源性Shh可以直接促進外周血管內皮細胞的管腔形成[21, 22];同時在胰腺癌、肺癌、前列腺癌和眼瞼腫瘤中也發現Shh的表達有不同程度增高[7, 9, 10, 12]。本研究結果顯示,血清饑餓24h后, 即刻可見hRPE細胞有少量Shh表達, 說明hRPE細胞在基礎情況下即有內源性Shh的表達。Cont組細胞中Shh mRNA呈現出低表達狀態,而CoCl2組細胞內Shh mRNA表達相對升高,并且隨缺氧時間的增加,Shh mRNA的表達呈現逐漸增加的趨勢。說明Shh的表達可以作為視網膜新生血管性疾病的一項監測指標,但具體通過何種方式監測有待進一步研究。同時本研究結果發現,CoCl2組細胞內VEGF mRNA的表達也相應提高,而CYA組細胞內VEGF mRNA的表達明顯降低。由此推斷,Shh可能參與了hRPE細胞中VEGF某條信號的傳導,從而減少VEGF的表達。具體Shh如何影響VEGF,以及在VEGF信號通路網絡中占據多大的作用也待進一步探究。
Cyclopamine是從藜蘆屬植物內分離得到的一種異甾體類生物堿,對Hh信號通路有抑制作用[23]。Cyclopamine從獨眼羊中發現[23],其對于人類視網膜細胞及組織的毒性及有效劑量仍是目前研究的焦點。既往對cyclopamine的毒性作用有過不同濃度的研究,發現20μmol/L的cyclopamine對hRPE細胞無明顯毒副作用。本研究結果顯示,體外條件下,20μmol/L的cyclopamine對缺氧誘導的hRPE細胞中VEGF的表達具有明顯的抑制作用。Cyclopamine抑制Shh信號途徑的作用及其對胚胎發育過程中異常信號的調控,也許可成為治療視網膜新生血管性疾病的有效方法。
缺氧刺激能夠顯著促進hRPE細胞的增生和分化[24]。在視網膜、脈絡膜新生血管疾病的發生和進展中, 視網膜、脈絡膜的缺血缺氧可以直接引起RPE細胞的一系列應激反應, 導致內皮細胞分裂和增生, 從而導致新生血管以及VEGF表達增多[25]。采用CoCl2處理細胞,可誘導細胞內對低氧敏感的基因表達增加。本研究所采用的造模方法應用成熟,已有研究者成功應用此方法制作缺氧模型[26]。應用CoCl2造成缺氧的原因之一可能是CoCl2阻礙脯氨酰羥化酶的活性,從而導致缺氧誘導因子-1α蛋白的積累,這與在缺氧培養箱中培養可達到大致相同的作用[27, 28]。本研究結果發現,Shh信號通路的激活與VEGF的表達相關, 提示Shh信號通路可能在缺氧誘導的hRPE細胞中VEGF表達具有一定作用。不足之處在于本研究實驗僅有細胞實驗結果,缺乏免疫組織化學或免疫熒光影像觀察內容,有待進一步觀察研究。
血管內皮生長因子(VEGF)為重要的促新生血管生成因子, 參與缺血缺氧誘導的視網膜、脈絡膜新生血管的形成[1-3]。視網膜周細胞、微血管內皮細胞、Müller細胞和視網膜色素上皮(RPE)細胞皆可產生VEGF, 其中以RPE細胞表達VEGF能力最為顯著[4]。Hedgehog (Hh)信號通路在調控脊椎動物細胞增生和血管發生、生成中起到重要作用[5]。脊椎動物Hh基因包含音猬因子(Shh)、沙漠刺猬因子(Dhh)、印度刺猬因子(Ihh),其中Shh信號與血管發育密切相關[6]。研究結果表明,眾多腫瘤的發生與Shh信號通路相關[7-12],并且已有臨床數據證實腫瘤組織中Shh信號通路與VEGF的表達存在相關性,但兩者具體作用機制尚不明確[13]。近期研究結果顯示, 缺氧可上調人RPE(hRPE)細胞中VEGF的表達[14-16],但其信號轉導機制尚不完全清楚。為此,本研究初步探討Shh信號轉導通路在缺氧誘導hRPE細胞VEGF表達中的作用。現將結果報道如下。
1 材料和方法
hRPE-19(ARPE-19)細胞株(中山大學動物實驗中心);Dulbecco改良Eagle(DMEM)培養基、青鏈霉素、胰蛋白酶[中科邁晨(北京)科技有限公司];胎牛血清(杭州四季青生物工程材料有限公司);cyclopamine、CoCl2(美國Sigma公司);Trizol試劑,逆轉錄(RT)試劑盒、熒光定量聚合酶鏈反應(RT-PCR)試劑盒(大連寶生物工程技術服務有限公司);人VEGF酶鏈免疫吸附測定試驗(ELISA)試劑盒(上海依科賽生物技術公司)。
ARPE-19細胞株接種于含10%胎牛血清、100×103U/ml青霉素、100 mg/ml鏈霉素的DMEM培養液中,置于37℃,5%CO2的培養箱中培養,每3~4天換液傳代,取對數生長期3~6代細胞用于實驗。以細胞密度1×104、10×105、5×105個/孔分別接種于96、6、24孔板中,培養24 h后棄上清液,每孔加入無血清DMEM培養液饑餓24 h,將細胞分為正常對照組(Cont組)、缺氧組。缺氧組加入100μmol/L CoCl2后再分為缺氧對照組(CoCl2組)、cyclopamine處理組(CYA組)、二甲基亞砜對照組(DMSO組)。僅CYA組于造成缺氧前1 h加入20μmol/L的cyclopamine進行預處理[17, 18];DMSO組加入1‰DMSO。各組細胞培養4、8、12、24 h。
RT-PCR檢測各組細胞中Shh、VEGF mRNA的表達。取培養4、8、12、24 h的6孔板,分別提取各組細胞總RNA,逆轉錄成cDNA后進行擴增,紫外分光光度計測定RNA濃度為3.5~5.0 ng/μl,吸光度[A,舊稱光密度(OD)]260/A280為1.8~2.0。按照RT試劑盒說明書操作,反應體系為10μl。Shh、VEGF引物序列由大連寶生物工程技術服務有限公司合成。Shh擴增片段長度94堿基對(bp),上游引物:5′-TGT CTGCTGCTAGTCCTCGTCTC-3′,下游引物:5′-TT TTGGGGTGCCTCCTCTT-3′;VEGF擴增片段長度116 bp,上游引物:5′-CTGGAGTGTGTGCCCACTG A-3′,下游引物:5′-CATTCACATTTGTTGTGCTG TAGGA-3′;β-肌動蛋白(β-actin)擴增片段長度186 bp,上游引物:5′-TGGCACCCAGCACAATGAA-3′,下游引物:5′-CTAAGTCATAGTCCGCCTAGAAGCA-3′。擴增條件:94℃預變性30 s,95℃變性5 s,65℃退火30 s,39個循環;65℃延伸10 s。按照2-ΔΔCt計算各組VEGF、Shh mRNA相對表達量。實驗重復3次,取平均值。
ELISA檢測細胞培養上清液中VEGF蛋白含量。培養4、8、12、24 h的24孔板ARPE-19細胞分為Cont組、CoCl2組、CYA組、DMSO組,每組設3個副孔。取細胞培養上清液, 離心半徑4.5 cm,6000 r/min離心5 min。采用雙抗體夾心ELISA法,按照人VEGF ELISA試劑盒說明書進行操作。酶標儀450 nm波長讀取A值, 待測樣本的A值在標準曲線上查出該樣本的VEGF水平對應值。實驗重復3次,取平均值。
應用SPSS 13.0統計軟件行統計學分析處理。數據以均數±標準差(
2 結果
RT-PCR檢測結果顯示,缺氧組細胞內Shh、VEGF mRNA表達隨時間延長而逐漸增高,與Cont組細胞內Shh、VEGF mRNA表達比較,差異均有統計學意義(F=45.260、264.938,P=0.001、0.001)。CYA組細胞內Shh、VEGF mRNA表達較CoCl2組降低,差異均有統計學意義(P<0.01);DMSO組細胞內Shh、VEGF mRNA表達與CoCl2組比較,差異均無統計學意義(P>0.05)(圖 1, 2)。
圖1
各組細胞內Shh mRNA表達比較
圖2
各組細胞內VEGF mRNA表達比較
ELISA檢測結果顯示,CoCl2組細胞培養上清液中VEGF蛋白含量隨時間延長而顯著增加,與Cont組細胞培養上清液中VEGF蛋白含量比較,差異均有統計學意義(F=3 156.676,P=0.001);CYA組細胞培養上清液中VEGF蛋白含量較CoCl2組明顯降低, 差異均有統計學意義(P<0.01)(圖 3)。
圖3
各組細胞培養上清液中VEGF蛋白含量比較
3 討論
Hh信號在血管發生和生成中起重要作用[19]。脊椎動物的Hh基因包含Shh、Dhh和Ihh[6],其中Shh信號與血管發育密切相關且對多種器官的形態發生分化不可或缺[19]。Shh被認為是一種脊椎動物體內間接的血管生成因子,可以直接作用于內皮細胞或刺激血管支持細胞產生上調VEGF和血管生成素[20]。既往研究發現,在生理條件下,外源性Shh可以直接促進外周血管內皮細胞的管腔形成[21, 22];同時在胰腺癌、肺癌、前列腺癌和眼瞼腫瘤中也發現Shh的表達有不同程度增高[7, 9, 10, 12]。本研究結果顯示,血清饑餓24h后, 即刻可見hRPE細胞有少量Shh表達, 說明hRPE細胞在基礎情況下即有內源性Shh的表達。Cont組細胞中Shh mRNA呈現出低表達狀態,而CoCl2組細胞內Shh mRNA表達相對升高,并且隨缺氧時間的增加,Shh mRNA的表達呈現逐漸增加的趨勢。說明Shh的表達可以作為視網膜新生血管性疾病的一項監測指標,但具體通過何種方式監測有待進一步研究。同時本研究結果發現,CoCl2組細胞內VEGF mRNA的表達也相應提高,而CYA組細胞內VEGF mRNA的表達明顯降低。由此推斷,Shh可能參與了hRPE細胞中VEGF某條信號的傳導,從而減少VEGF的表達。具體Shh如何影響VEGF,以及在VEGF信號通路網絡中占據多大的作用也待進一步探究。
Cyclopamine是從藜蘆屬植物內分離得到的一種異甾體類生物堿,對Hh信號通路有抑制作用[23]。Cyclopamine從獨眼羊中發現[23],其對于人類視網膜細胞及組織的毒性及有效劑量仍是目前研究的焦點。既往對cyclopamine的毒性作用有過不同濃度的研究,發現20μmol/L的cyclopamine對hRPE細胞無明顯毒副作用。本研究結果顯示,體外條件下,20μmol/L的cyclopamine對缺氧誘導的hRPE細胞中VEGF的表達具有明顯的抑制作用。Cyclopamine抑制Shh信號途徑的作用及其對胚胎發育過程中異常信號的調控,也許可成為治療視網膜新生血管性疾病的有效方法。
缺氧刺激能夠顯著促進hRPE細胞的增生和分化[24]。在視網膜、脈絡膜新生血管疾病的發生和進展中, 視網膜、脈絡膜的缺血缺氧可以直接引起RPE細胞的一系列應激反應, 導致內皮細胞分裂和增生, 從而導致新生血管以及VEGF表達增多[25]。采用CoCl2處理細胞,可誘導細胞內對低氧敏感的基因表達增加。本研究所采用的造模方法應用成熟,已有研究者成功應用此方法制作缺氧模型[26]。應用CoCl2造成缺氧的原因之一可能是CoCl2阻礙脯氨酰羥化酶的活性,從而導致缺氧誘導因子-1α蛋白的積累,這與在缺氧培養箱中培養可達到大致相同的作用[27, 28]。本研究結果發現,Shh信號通路的激活與VEGF的表達相關, 提示Shh信號通路可能在缺氧誘導的hRPE細胞中VEGF表達具有一定作用。不足之處在于本研究實驗僅有細胞實驗結果,缺乏免疫組織化學或免疫熒光影像觀察內容,有待進一步觀察研究。