引用本文: 胡健艷, 李婷婷, 吳強. G蛋白偶聯受體91對早期糖尿病大鼠血視網膜屏障的影響及機制. 中華眼底病雜志, 2015, 31(2): 157-161. doi: 10.3760/cma.j.issn.1005-1015.2015.02.012 復制
糖尿病視網膜病變(DR)是由于長期慢性高血糖引起視網膜血管內皮細胞損傷、神經細胞凋亡及視網膜色素上皮(RPE)細胞功能障礙,導致血視網膜屏障(BRB)破環而出現的滲出、水腫及出血等一系列病理改變。血管內皮生長因子(VEGF)是DR發生發展過程中的重要調節因子,其表達增加與視網膜BRB損傷密切相關[1-3]。G蛋白偶聯受體91(GPR91)主要表達在肝臟、脾臟、胎盤、腎臟及視網膜等血供豐富的組織,具有調節血管生長以適應組織代謝需要的作用[4]。我們的前期研究結果顯示,高糖環境下激活的GPR91參與調控VEGF的表達和分泌[5-7]。為進一步探討GPR91在DR中的可能作用,我們通過玻璃體腔注射攜帶GPR91小發夾RNA(shRNA)慢病毒載體, 觀察了早期糖尿病大鼠視網膜中GPR91對VEGF表達的調控及視網膜BRB損傷的影響及可能的機制。現將結果報道如下。
1 材料和方法
鏈脲佐菌素(STZ)(美國Sigma公司);GPR91抗體(美國Novus Biologicals公司),絲裂原活化蛋白激酶(MAPK)通路抗體及磷酸化抗體:t-細胞外信號調節激酶(ERK)1/2、t-c-jun氨基末端激酶(JNK)、t-p38 MAPK、p-ERK1/2、p-JNK、p-p38 MAPK(美國Cell Signalling公司),β-肌動蛋白抗體(北京康為公司),二抗(美國ProteinTech Group公司);VEGF酶聯免疫吸附試驗(ELISA)試劑盒(美國R & D公司);血糖儀、血糖試紙(美國雅培公司),微量注射器(德國Hamilton公司),凝膠成像分析儀(美國Bio-Rad公司),多功能酶標儀(美國MD公司),光學顯微鏡(德國Leica公司)。慢病毒pGCSIL-GFP-shGPR91及相應的空載體構建方法同文獻[7]。
健康成年雄性Sprague-Dawley大鼠60只,鼠齡8~9周,體重200~250 g,清潔級,上海市第六人民醫院動物實驗中心提供。喂養環境控制室溫在23~25℃,相對濕度50%~60%;自由攝食、飲水。采用隨機數字表法將大鼠分為對照組(A組)、糖尿病STZ組(B組)、空病毒LV.shScrambled組(C組)、病毒LV.shGPR91組(D組),每組均為15只大鼠。除A組大鼠,其他各組大鼠參照文獻[6]的方法,按體重60 mg/kg腹腔注射STZ制作糖尿病模型。成模2周,C組大鼠玻璃體腔注射濃度為1×108 TU/ml的空載體病毒1 μl;D組大鼠玻璃體腔注射濃度為1×108 TU/ml的pGCSIL-GFP-shGPR91慢病毒1 μl。TU/ml=觀察到帶有熒光的細胞個數/病毒原液量[7]。各組從玻璃體腔注射載體病毒后開始計算,觀察12周。
大鼠過量麻醉處死,迅速摘除眼球,4%多聚甲醛固定24 h,常規脫水,石蠟包埋,作5μm厚度的連續切片。采用蘇木精-伊紅(HE)染色觀察視網膜微血管的病理改變。采用免疫組織化學染色法檢測GPR91的表達。脫蠟和水化后,3%過氧化氫消除內源性過氧化物酶,正常山羊血清封閉,兔抗大鼠GPR91單克隆抗體1∶500稀釋,4℃過夜,生物素標記的羊抗兔抗體室溫條件下孵育60 min,3,3′-二氨基聯苯胺(DAB)顯色,蘇木精復染,封片,光學顯微鏡下觀察。
視網膜通透性檢測,評價BRB損傷。參照文獻[8]的方法,伊凡思藍(EB)按體重45 mg/kg由股靜脈注入。循環2 h后,37℃預熱的0.05 mmol/L檸檬酸緩沖液(pH3.5)心臟灌注。立即摘除眼球,抽干視網膜,加甲酰胺70℃孵育18 h,分別測定620、740 nm的吸光度[A,舊稱光密度(OD)]。計算EB經視網膜血管的滲漏量:[視網膜EB量(mg)/視網膜干重(g)]/[單位時間內血漿EB量(mg)/血漿體積(μl)×循環時間(h)]。
蛋白免疫印跡法(Western blot)檢測視網膜GPR91、MAPK通路各分子的表達及激活。各組大鼠視網膜組織加入蛋白質裂解液提取蛋白,二喹啉甲酸法測定總蛋白濃度。取30 μg總蛋白,行十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳分離后,濕轉法將蛋白轉移至聚偏二氟乙烯膜。5%脫脂牛奶粉封閉1 h,分別加入GPR91單克隆抗體(1∶1000),p-ERK1/2多克隆抗體(1∶3000),t-ERK1/2多克隆抗體(1∶3000),p-JNK多克隆抗體(1∶1000),t-JNK多克隆抗體(1∶1000),p-p38 MAPK多克隆抗體(1∶1000)和t-p38 MAPK多克隆抗體(1∶1000)4℃過夜。加入辣根過氧化物酶標記的二抗(1∶5000),室溫孵育l h。增強化學發光法顯影,凝膠成像系統攝取圖像并進行分析。
采用ELISA測定VEGF蛋白含量。抽取各組大鼠的玻璃體液,4℃, 離心半徑5 cm,7600 r/min離心20 min, 取上清,按照VEGF試劑盒說明書進行測定。
SPSS 16.0統計學軟件進行統計分析。所有數據以均數±標準差(
2 結果
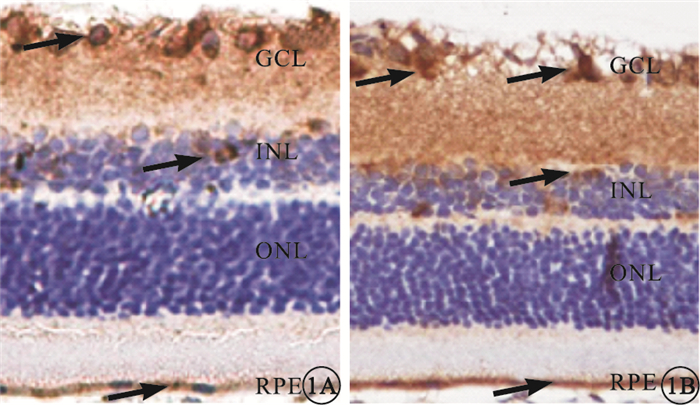
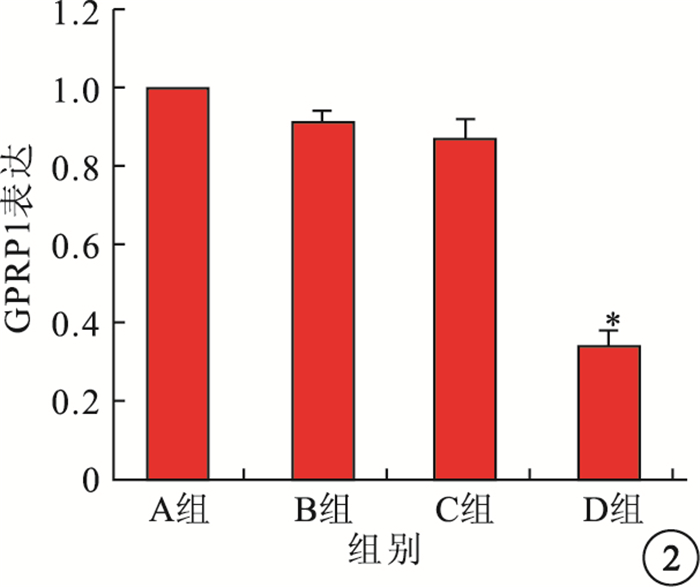
免疫組織化學染色可見,GPR91主要表達于大鼠視網膜神經節細胞層(GCL),小部分表達于內核層(INL)和RPE層(圖 1),外核層(ONL)未見其表達。Western blot檢測結果顯示,A、B、C組間GPR91表達比較,差異無統計學意義(F=1.69,P>0.05);D組GPR91表達較C組顯著降低,差異有統計學意義(F=39.31,P<0.01)(圖 2)。
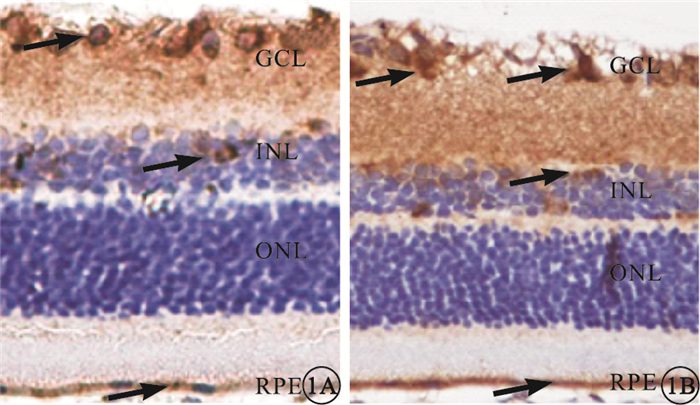 圖1
大鼠視網膜免疫組織化學染色像。1A.A組;1B.B組。GPR91主要表達于大鼠視網膜GCL,小部分表達于INL和RPE層(黑箭) DAB×200
圖1
大鼠視網膜免疫組織化學染色像。1A.A組;1B.B組。GPR91主要表達于大鼠視網膜GCL,小部分表達于INL和RPE層(黑箭) DAB×200
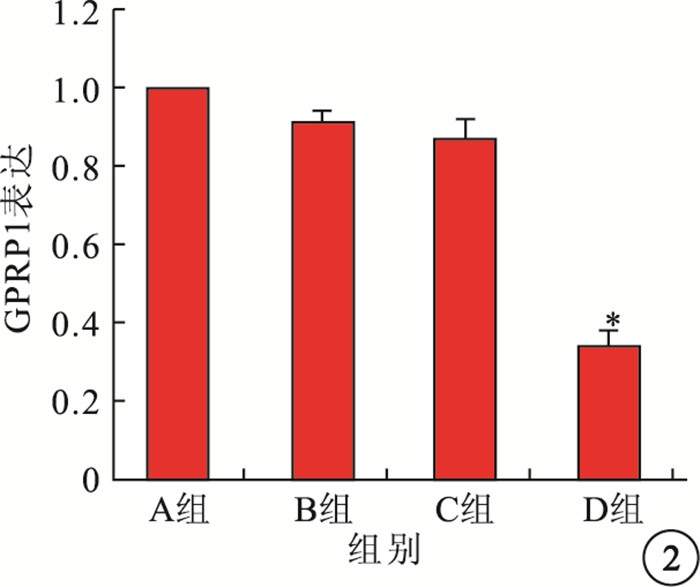 圖2
各組大鼠視網膜GPR91表達結果定量分析。*與C組比較,P<0.01
圖2
各組大鼠視網膜GPR91表達結果定量分析。*與C組比較,P<0.01
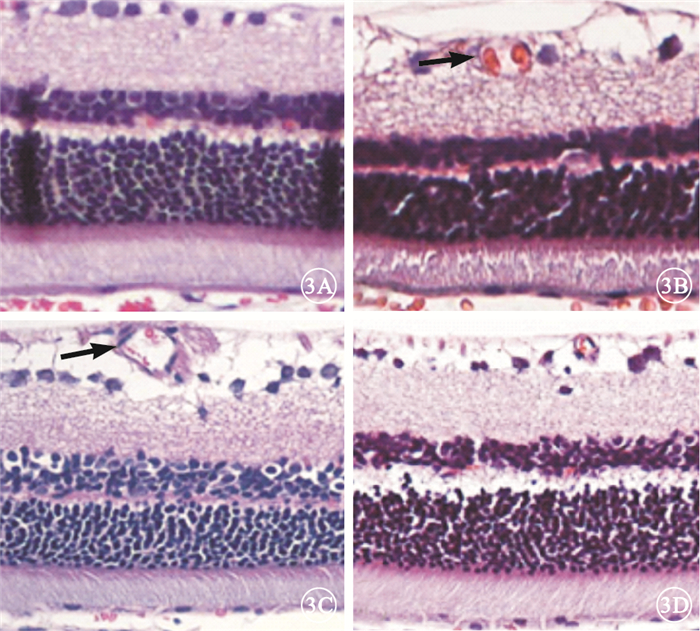
光學顯微鏡觀察發現,B、C組大鼠視網膜內層毛細血管較A組大鼠視網膜內層毛細血管明顯擴張,D組大鼠視網膜內層擴張的毛細血管顯著改善(圖 3)。
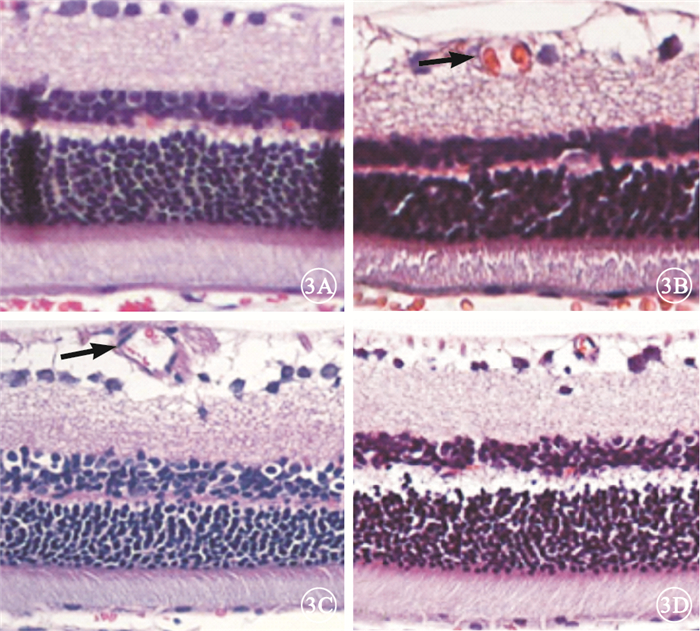 圖3
各組大鼠視網膜光學顯微鏡像。3A~3D.分別為A~D組。B、C組大鼠視網膜內層毛細血管較A組大鼠視網膜內層毛細血管明顯擴張(黑箭),而D組大鼠視網膜內層毛細血管擴張改善HE×200
圖3
各組大鼠視網膜光學顯微鏡像。3A~3D.分別為A~D組。B、C組大鼠視網膜內層毛細血管較A組大鼠視網膜內層毛細血管明顯擴張(黑箭),而D組大鼠視網膜內層毛細血管擴張改善HE×200
視網膜通透性檢測結果顯示, B、C組大鼠視網膜EB滲漏量分別是A組的(1.58±0.11)、(1.48±0.10)倍,D組大鼠視網膜EB滲漏量較C組下降(33.8±4.11)%,差異有統計學意義(F=30.35,P<0.05)。
ELISA結果顯示,A~D組大鼠視網膜VEGF蛋白表達量分別為(58.44±16.17)、(453.87±24.36)、(409.94±22.16)、(77.32±18.1) pg/ml。B、C組大鼠視網膜VEGF蛋白表達較A組明顯增加;D組大鼠視網膜VEGF蛋白表達較C組顯著下降,差異有統計學意義(F=253.15,P<0.05)。
Western blot檢測結果顯示,B組ERK1/2、JNK、p38 MAPK通路均激活, B組p-ERK1/2/t-ERK1/2、p-JNK/t-JNK、p-p38 MAPK/t-p38 MAPK比值與A組比較,差異均有統計學意義(q=6.38、2.94、3.45,P<0.05)。C組各比值與B組比較,差異無統計學意義(q=-0.60、-0.35、-0.68,P>0.05);D組中p-ERK1/2與t-ERK1/2的比值較C組顯著降低,差異有統計學意義(F=22.50,P<0.05);p-JNK/t-JNK和p-p38 MAPK/t-p38 MAPK比值未見明顯下調,差異無統計學意義(F=4.76、4.57,P>0.05)(表 1)。
3 討論
本研究通過單次腹腔注射STZ的方式建立糖尿病大鼠模型,12周后觀察發現早期糖尿病大鼠視網膜內層血管明顯擴張,BRB被破壞,血管通透性增加,EB滲漏顯著增加,與Zhang和Yan[9]的研究結果相一致。當玻璃體腔內轉入攜帶有GPR91 shRNA的慢病毒載體時,糖尿病大鼠視網膜血管擴張程度改善,BRB破壞程度減輕,血管滲漏量顯著減少。GPR91受體又稱琥珀酸受體1,屬于G蛋白偶聯受體家族,具有7次跨膜螺旋的結構特點,通過特異性結合相應配體琥珀酸參與多種生理及病理反應[10]。Matsumoto等[11]研究發現,增生型DR患者的玻璃體腔內琥珀酸含量顯著增加。我們的前期研究通過氣相色譜質譜儀分析觀察到糖尿病大鼠視網膜中也存在著琥珀酸蓄積現象[6]。因此,我們推測糖尿病環境下,隨著視網膜中琥珀酸的異常增多,激活GPR91受體參與DR引起的BRB破壞。
目前已有大量研究證實,VEGF的異常增多可破壞視網膜血管內皮細胞間的緊密連接,促進內皮細胞的增生與遷移,最終導致新生血管形成[12, 13]。本研究也觀察到糖尿病大鼠視網膜分泌的VEGF蛋白較血糖正常大鼠明顯增加,玻璃體腔內注射攜帶有GPR91 shRNA的慢病毒載體后可明顯減弱VEGF的分泌。Sapieha等[4]研究發現,缺氧條件下視網膜神經節細胞中GPR91蛋白的激活可引起VEGF的表達增加并誘導視網膜新生血管的形成,但去除GCL后VEGF水平顯著下調。因此,我們認為GPR91對BRB功能的調控可能是通過調節VEGF的分泌實現的。
為了進一步研究GPR91調控VEGF的信號通路,我們探討MAPK通路在其發揮中的作用。MAPK是一種絲氨酸/蘇氨酸蛋白激酶,可被生長因子、細胞因子、激素、神經遞質等廣泛刺激激活,是生物體內重要的信號轉導系統之一[14, 15]。大量研究結果表明,MAPK信號通路在眼底新生血管發生發展中也起著重要調控作用[16, 17]。本研究發現,早期糖尿病大鼠視網膜中MARK信號通路3個成員(ERK、JNK、p38 MAPK)均激活,同時ERK1/2信號通路參與GPR91調控作用,結果與Vargas等[18]在糖尿病腎病中的研究結果相似。ERK1/2信號通路在促進蛋白質合成、細胞增生、腫瘤形成等方面發揮重要作用[19]。近幾年,隨著對DR發病機制研究的增加,ERK1/2信號通路在DR中調節VEGF表達方面的相關報道也越來越多[20, 21]。結合本研究結果,我們推測GPR91對VEGF的調控作用可能通過ERK1/2信號通路實現,其具體調控機制將是下一步研究重點。
本研究結果表明,GPR91受體在DR視網膜BRB功能受損和VEGF的分泌表達中發揮著重要調控作用,提示GPR91基因將有可能成為有效控制DR的重要位點之一。由于DR是多種分子和信號通路相互作用的復雜過程,因此我們不能完全排除其他機制參與的可能。
糖尿病視網膜病變(DR)是由于長期慢性高血糖引起視網膜血管內皮細胞損傷、神經細胞凋亡及視網膜色素上皮(RPE)細胞功能障礙,導致血視網膜屏障(BRB)破環而出現的滲出、水腫及出血等一系列病理改變。血管內皮生長因子(VEGF)是DR發生發展過程中的重要調節因子,其表達增加與視網膜BRB損傷密切相關[1-3]。G蛋白偶聯受體91(GPR91)主要表達在肝臟、脾臟、胎盤、腎臟及視網膜等血供豐富的組織,具有調節血管生長以適應組織代謝需要的作用[4]。我們的前期研究結果顯示,高糖環境下激活的GPR91參與調控VEGF的表達和分泌[5-7]。為進一步探討GPR91在DR中的可能作用,我們通過玻璃體腔注射攜帶GPR91小發夾RNA(shRNA)慢病毒載體, 觀察了早期糖尿病大鼠視網膜中GPR91對VEGF表達的調控及視網膜BRB損傷的影響及可能的機制。現將結果報道如下。
1 材料和方法
鏈脲佐菌素(STZ)(美國Sigma公司);GPR91抗體(美國Novus Biologicals公司),絲裂原活化蛋白激酶(MAPK)通路抗體及磷酸化抗體:t-細胞外信號調節激酶(ERK)1/2、t-c-jun氨基末端激酶(JNK)、t-p38 MAPK、p-ERK1/2、p-JNK、p-p38 MAPK(美國Cell Signalling公司),β-肌動蛋白抗體(北京康為公司),二抗(美國ProteinTech Group公司);VEGF酶聯免疫吸附試驗(ELISA)試劑盒(美國R & D公司);血糖儀、血糖試紙(美國雅培公司),微量注射器(德國Hamilton公司),凝膠成像分析儀(美國Bio-Rad公司),多功能酶標儀(美國MD公司),光學顯微鏡(德國Leica公司)。慢病毒pGCSIL-GFP-shGPR91及相應的空載體構建方法同文獻[7]。
健康成年雄性Sprague-Dawley大鼠60只,鼠齡8~9周,體重200~250 g,清潔級,上海市第六人民醫院動物實驗中心提供。喂養環境控制室溫在23~25℃,相對濕度50%~60%;自由攝食、飲水。采用隨機數字表法將大鼠分為對照組(A組)、糖尿病STZ組(B組)、空病毒LV.shScrambled組(C組)、病毒LV.shGPR91組(D組),每組均為15只大鼠。除A組大鼠,其他各組大鼠參照文獻[6]的方法,按體重60 mg/kg腹腔注射STZ制作糖尿病模型。成模2周,C組大鼠玻璃體腔注射濃度為1×108 TU/ml的空載體病毒1 μl;D組大鼠玻璃體腔注射濃度為1×108 TU/ml的pGCSIL-GFP-shGPR91慢病毒1 μl。TU/ml=觀察到帶有熒光的細胞個數/病毒原液量[7]。各組從玻璃體腔注射載體病毒后開始計算,觀察12周。
大鼠過量麻醉處死,迅速摘除眼球,4%多聚甲醛固定24 h,常規脫水,石蠟包埋,作5μm厚度的連續切片。采用蘇木精-伊紅(HE)染色觀察視網膜微血管的病理改變。采用免疫組織化學染色法檢測GPR91的表達。脫蠟和水化后,3%過氧化氫消除內源性過氧化物酶,正常山羊血清封閉,兔抗大鼠GPR91單克隆抗體1∶500稀釋,4℃過夜,生物素標記的羊抗兔抗體室溫條件下孵育60 min,3,3′-二氨基聯苯胺(DAB)顯色,蘇木精復染,封片,光學顯微鏡下觀察。
視網膜通透性檢測,評價BRB損傷。參照文獻[8]的方法,伊凡思藍(EB)按體重45 mg/kg由股靜脈注入。循環2 h后,37℃預熱的0.05 mmol/L檸檬酸緩沖液(pH3.5)心臟灌注。立即摘除眼球,抽干視網膜,加甲酰胺70℃孵育18 h,分別測定620、740 nm的吸光度[A,舊稱光密度(OD)]。計算EB經視網膜血管的滲漏量:[視網膜EB量(mg)/視網膜干重(g)]/[單位時間內血漿EB量(mg)/血漿體積(μl)×循環時間(h)]。
蛋白免疫印跡法(Western blot)檢測視網膜GPR91、MAPK通路各分子的表達及激活。各組大鼠視網膜組織加入蛋白質裂解液提取蛋白,二喹啉甲酸法測定總蛋白濃度。取30 μg總蛋白,行十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳分離后,濕轉法將蛋白轉移至聚偏二氟乙烯膜。5%脫脂牛奶粉封閉1 h,分別加入GPR91單克隆抗體(1∶1000),p-ERK1/2多克隆抗體(1∶3000),t-ERK1/2多克隆抗體(1∶3000),p-JNK多克隆抗體(1∶1000),t-JNK多克隆抗體(1∶1000),p-p38 MAPK多克隆抗體(1∶1000)和t-p38 MAPK多克隆抗體(1∶1000)4℃過夜。加入辣根過氧化物酶標記的二抗(1∶5000),室溫孵育l h。增強化學發光法顯影,凝膠成像系統攝取圖像并進行分析。
采用ELISA測定VEGF蛋白含量。抽取各組大鼠的玻璃體液,4℃, 離心半徑5 cm,7600 r/min離心20 min, 取上清,按照VEGF試劑盒說明書進行測定。
SPSS 16.0統計學軟件進行統計分析。所有數據以均數±標準差(
2 結果
免疫組織化學染色可見,GPR91主要表達于大鼠視網膜神經節細胞層(GCL),小部分表達于內核層(INL)和RPE層(圖 1),外核層(ONL)未見其表達。Western blot檢測結果顯示,A、B、C組間GPR91表達比較,差異無統計學意義(F=1.69,P>0.05);D組GPR91表達較C組顯著降低,差異有統計學意義(F=39.31,P<0.01)(圖 2)。
圖1
大鼠視網膜免疫組織化學染色像。1A.A組;1B.B組。GPR91主要表達于大鼠視網膜GCL,小部分表達于INL和RPE層(黑箭) DAB×200
圖2
各組大鼠視網膜GPR91表達結果定量分析。*與C組比較,P<0.01
光學顯微鏡觀察發現,B、C組大鼠視網膜內層毛細血管較A組大鼠視網膜內層毛細血管明顯擴張,D組大鼠視網膜內層擴張的毛細血管顯著改善(圖 3)。
圖3
各組大鼠視網膜光學顯微鏡像。3A~3D.分別為A~D組。B、C組大鼠視網膜內層毛細血管較A組大鼠視網膜內層毛細血管明顯擴張(黑箭),而D組大鼠視網膜內層毛細血管擴張改善HE×200
視網膜通透性檢測結果顯示, B、C組大鼠視網膜EB滲漏量分別是A組的(1.58±0.11)、(1.48±0.10)倍,D組大鼠視網膜EB滲漏量較C組下降(33.8±4.11)%,差異有統計學意義(F=30.35,P<0.05)。
ELISA結果顯示,A~D組大鼠視網膜VEGF蛋白表達量分別為(58.44±16.17)、(453.87±24.36)、(409.94±22.16)、(77.32±18.1) pg/ml。B、C組大鼠視網膜VEGF蛋白表達較A組明顯增加;D組大鼠視網膜VEGF蛋白表達較C組顯著下降,差異有統計學意義(F=253.15,P<0.05)。
Western blot檢測結果顯示,B組ERK1/2、JNK、p38 MAPK通路均激活, B組p-ERK1/2/t-ERK1/2、p-JNK/t-JNK、p-p38 MAPK/t-p38 MAPK比值與A組比較,差異均有統計學意義(q=6.38、2.94、3.45,P<0.05)。C組各比值與B組比較,差異無統計學意義(q=-0.60、-0.35、-0.68,P>0.05);D組中p-ERK1/2與t-ERK1/2的比值較C組顯著降低,差異有統計學意義(F=22.50,P<0.05);p-JNK/t-JNK和p-p38 MAPK/t-p38 MAPK比值未見明顯下調,差異無統計學意義(F=4.76、4.57,P>0.05)(表 1)。
3 討論
本研究通過單次腹腔注射STZ的方式建立糖尿病大鼠模型,12周后觀察發現早期糖尿病大鼠視網膜內層血管明顯擴張,BRB被破壞,血管通透性增加,EB滲漏顯著增加,與Zhang和Yan[9]的研究結果相一致。當玻璃體腔內轉入攜帶有GPR91 shRNA的慢病毒載體時,糖尿病大鼠視網膜血管擴張程度改善,BRB破壞程度減輕,血管滲漏量顯著減少。GPR91受體又稱琥珀酸受體1,屬于G蛋白偶聯受體家族,具有7次跨膜螺旋的結構特點,通過特異性結合相應配體琥珀酸參與多種生理及病理反應[10]。Matsumoto等[11]研究發現,增生型DR患者的玻璃體腔內琥珀酸含量顯著增加。我們的前期研究通過氣相色譜質譜儀分析觀察到糖尿病大鼠視網膜中也存在著琥珀酸蓄積現象[6]。因此,我們推測糖尿病環境下,隨著視網膜中琥珀酸的異常增多,激活GPR91受體參與DR引起的BRB破壞。
目前已有大量研究證實,VEGF的異常增多可破壞視網膜血管內皮細胞間的緊密連接,促進內皮細胞的增生與遷移,最終導致新生血管形成[12, 13]。本研究也觀察到糖尿病大鼠視網膜分泌的VEGF蛋白較血糖正常大鼠明顯增加,玻璃體腔內注射攜帶有GPR91 shRNA的慢病毒載體后可明顯減弱VEGF的分泌。Sapieha等[4]研究發現,缺氧條件下視網膜神經節細胞中GPR91蛋白的激活可引起VEGF的表達增加并誘導視網膜新生血管的形成,但去除GCL后VEGF水平顯著下調。因此,我們認為GPR91對BRB功能的調控可能是通過調節VEGF的分泌實現的。
為了進一步研究GPR91調控VEGF的信號通路,我們探討MAPK通路在其發揮中的作用。MAPK是一種絲氨酸/蘇氨酸蛋白激酶,可被生長因子、細胞因子、激素、神經遞質等廣泛刺激激活,是生物體內重要的信號轉導系統之一[14, 15]。大量研究結果表明,MAPK信號通路在眼底新生血管發生發展中也起著重要調控作用[16, 17]。本研究發現,早期糖尿病大鼠視網膜中MARK信號通路3個成員(ERK、JNK、p38 MAPK)均激活,同時ERK1/2信號通路參與GPR91調控作用,結果與Vargas等[18]在糖尿病腎病中的研究結果相似。ERK1/2信號通路在促進蛋白質合成、細胞增生、腫瘤形成等方面發揮重要作用[19]。近幾年,隨著對DR發病機制研究的增加,ERK1/2信號通路在DR中調節VEGF表達方面的相關報道也越來越多[20, 21]。結合本研究結果,我們推測GPR91對VEGF的調控作用可能通過ERK1/2信號通路實現,其具體調控機制將是下一步研究重點。
本研究結果表明,GPR91受體在DR視網膜BRB功能受損和VEGF的分泌表達中發揮著重要調控作用,提示GPR91基因將有可能成為有效控制DR的重要位點之一。由于DR是多種分子和信號通路相互作用的復雜過程,因此我們不能完全排除其他機制參與的可能。