引用本文: 薄其玉, 東莉潔, 張琰, 周玉, 劉勛, 韓倩, 王飛. 氧誘導視網膜病變小鼠模型微小RNA表達譜分析. 中華眼底病雜志, 2015, 31(1): 77-82. doi: 10.3760/cma.j.issn.1005-1015.2015.01.019 復制
玻璃體腔注射抗血管內皮生長因子(VEGF)藥物是目前治療眼部新生血管性疾病的有效方法,但其存在并發癥、副作用多等問題,甚至部分患者經抗VEGF藥物治療后仍不能阻止視網膜新生血管(RNV)的形成[1-3]。尋找不依賴VEGF的促新生血管生成的其他通路,以及高效、低損傷拮抗RNV的治療靶點尤為重要。微小RNA由具有發夾結構的單鏈RNA前體經Dicer酶加工后生成,主要通過對靶mRNA的降解或翻譯抑制,使特定靶基因沉默而發揮作用[4]。目前已明確微小RNA在眼部組織發育分化、細胞凋亡、代謝和損傷后再生等方面具有調控作用[5]。但其是否在RNV形成過程中發揮作用還不清楚。為此,我們通過建立氧誘導視網膜病變(OIR)小鼠模型,利用微小RNA芯片分析技術檢測出具有差異表達的微小RNA并進行實時聚合酶鏈反應(RT-PCR)驗證,以探索與RNV形成有關的微小RNA。現將結果報道如下。
1 材料和方法
7日齡健康清潔級C57BL/6J小鼠52只,雌雄不限,由中國人民解放軍軍事醫學科學院實驗動物中心提供。按隨機數字表法將小鼠分為正常對照組和OIR組,每組26只。參照文獻[6]的方法建立OIR動物模型,將OIR組小鼠與哺乳母鼠一同置入密閉玻璃容器中,氣體流量控制在2 L/min左右,箱內含氧體積分數為(75±2)%;用CY-12C數字測氧儀(浙江建德梅城電化分析儀器廠)間斷性監測出氣口處氧濃度,6~8次/d。每日打開密閉容器1次,更換墊料、加食水,替換母鼠。高氧環境下連續飼養5 d后,再置于正常空氣中飼養5 d。正常對照組小鼠在正常空氣中飼養10 d,不做任何處理。
小鼠17日齡時,每組選取8只小鼠作視網膜鋪片,觀察小鼠視網膜血管形態,測量視網膜無灌注區的相對面積。參照文獻[7]的方法,取25 mg相對分子質量為2×106的異硫氰酸熒光素-葡萄聚糖(FITC-dextran)粉末(美國Sigma公司)溶于0.5 ml去離子水中,以離心半徑5.13 cm,13 000 r/min離心5 min,取上清液,制備50 mg/ml的FITC-dextran灌注液。以左手拇指及中指捏住小鼠頭頸部皮膚,食指輕壓眼眶使眼球突出,然后將依據小鼠眼球大小彎成弧形的1 ml注射器針頭刺入左眼內側眼角的Tenon氏筋膜,沿鼻側眼眶壁平行地對喉頭方向推進約2 mm達球后靜脈叢,略轉動注射器并去除眼后部壓力,緩慢注入0.05 ml的FITC-dextran灌注液。靜置5 min,頸椎脫臼法處死小鼠,摘除眼球置于4%的多聚甲醛液中固定40 min,之后放入磷酸鹽緩沖液中4℃避光保存。手術顯微鏡下,沿角鞏膜緣剪開眼球,去除角膜、虹膜,保留晶狀體在眼杯中。兩把鑷子配合牽拉眼杯后極部殘留的視神經,使鞏膜和脈絡膜與視網膜完整分離。去除晶狀體,將分離的視網膜以視盤為中心,放射狀切開4片,玻璃體面向上平鋪于載玻片上,用含4′, 6-二脒基-2-苯基吲哚(DAPI)的抗淬滅封片劑(美國Vectashield公司)封片,蓋上蓋玻片。熒光顯微鏡(日本Olympus公司)下觀察并照相。采用CellSens Standard軟件(日本Olympus公司)測量視網膜無灌注區占全視網膜的相對面積。
小鼠17日齡時,每組選取8只小鼠作視網膜石蠟切片,蘇木精-伊紅(HE)染色并計數突破視網膜內界膜的視網膜血管細胞核數。取小鼠眼球,常規方法制作蠟塊,行矢狀位3μm厚連續切片,相鄰2張切片間隔40μm,每只眼球取10張代表性切片行HE染色,排除含視神經的切片。統計平均每只眼球每張切片中突破內界膜的視網膜血管細胞核數。參照文獻[8, 9]的計數標準,僅計數與視網膜有緊密聯系的血管細胞核,不包括玻璃體腔內與內界膜無聯系的血管細胞核。
小鼠17日齡時,每組選取10只小鼠處死并取眼球,立即放于液氮中,并轉存于-80℃冰箱中。其中5只小鼠的眼球標本由廣州復能基因有限公司對80個微小RNA行芯片分析。另外5只小鼠的眼球在取得微小RNA芯片結果后采用RT-PCR驗證芯片檢測差異表達倍數≥3.0的微小RNA。采用E.Z.N.A.微小RNA Kit提取試劑盒(美國Omega Bio-Tek公司)提取微小RNA。將眼球組織放入預冷的玻璃管中研磨,并加入1 ml裂解液,冰上勻漿3 min,移至1.5 ml離心管中,室溫下孵育3 min。1 ml裂解液中加入200μl氯仿,蓋好管蓋后劇烈震蕩混勻,冰中孵育10 min, 以12 000×g在4℃條件下離心15 min。取上清液,并加入無水乙醇混勻,經試劑盒中的HiBind RNA Mini柱過濾去除mRNA,濾液再經過MicroElute RNA柱,使微小RNA吸附于過濾柱的濾膜上,最后用30μl洗脫液洗脫。Nanodrop 2000分光光度計測定microRNAs的含量和純度,-80℃保存待用。All-in-One? miRNA qRT-PCR Detection Kit試劑盒(Cat.No.AOMD-Q050,美國GeneCopoeia公司)逆轉微小RNA并進行PCR反應。取500 ng微小RNA,按說明書中的反應體系和操作步驟進行逆轉。根據微小RNA的濃度計算加入微小RNA的體積,并向離心管中依次加入微小RNA、PolyA聚合酶、逆轉錄酶、5倍反應試劑,并用滅菌水補足至25μl,渦旋混勻并離心后使用GeneAmp PCR System 2400擴增儀進行逆轉錄。參數:37℃孵育60 min,85℃變性5 min。合成的cDNA于-80℃保存備用。cDNA經5倍稀釋后與上、下游引物一起進行RT-PCR反應,其中以RNU6為內參基因。引物均購自廣州復能基因有限公司,分別為miR-3078(Cat#MmiRQP1700)、miR-140(Cat#MmiRQP0181)、miR-29b(Cat#MmiRQP0373)、miR-29c(Cat#MmiRQP0375)和RNU6(Cat#MmiRQP9002)。RT-PCR采用7900HT熒光定量PCR儀。擴增參數:50℃孵育2 min,95℃變性10 min,然后40次循環;95℃變性15 s,60℃退火和延伸1 min;并加入解離階段:95℃變性15 s,60℃退火15 s,然后95℃反應15 s,以判定擴增產物的特異性。擴增后,獲得各基因擴增的循環閾值(Ct),以2-ΔΔCt法分析所得數據。ΔCt=目的基因Ct-內參基因Ct,ΔCt=OIR組目的基因2ΔCt-正常對照組目的基因ΔCt。MicroRNA表達差異倍數用2-ΔΔCt計算,并以表達差異倍數的log2值作圖。
采用SPSS 20.0統計軟件行統計學分析處理。各組量化的實驗數據資料經Shapiro-Wilk檢驗呈正態分布,以均數±標準差(
2 結果
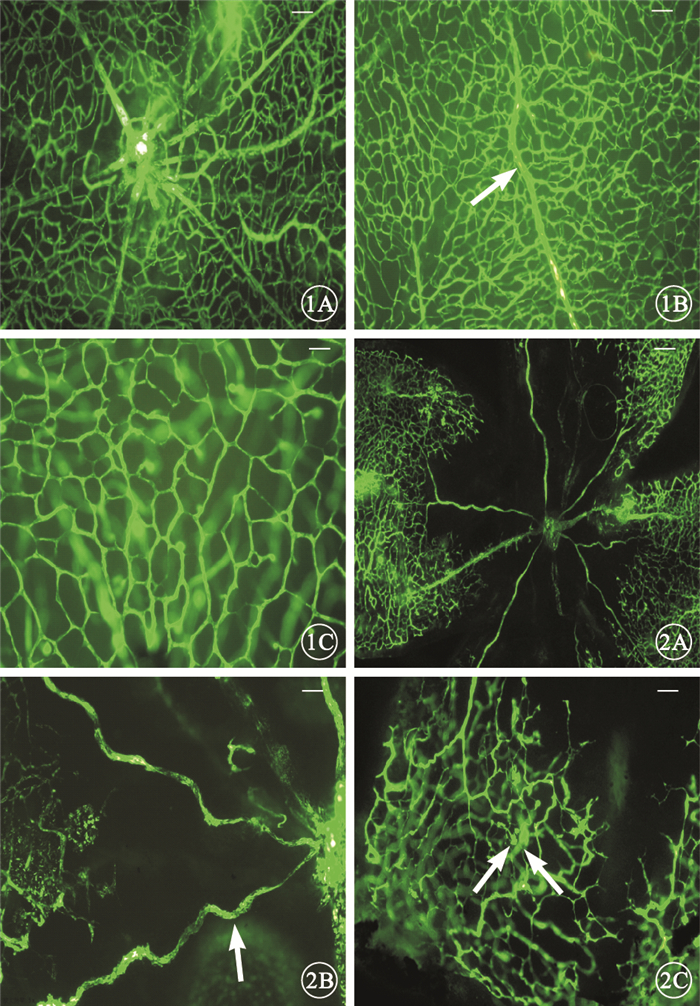
熒光顯微鏡觀察發現,正常對照組小鼠自視盤發出的大血管向四周呈放射狀均勻分布,直至視網膜周邊部,視網膜血管分布呈均勻的網狀結構(圖 1)。OIR組小鼠自視盤發出的大血管明顯紆曲,分支減少;中央可見大片無灌注區,中周部可見明顯的視網膜血管無灌注區和灌注區的分界,分界處可見大量的新生血管和新生血管芽,新生血管網結構及分布紊亂(圖 2)。定量分析結果顯示,正常對照組、OIR組小鼠視網膜無灌注區的相對面積分別為(6.57±3.6)%、(25.81±2.12)%。OIR組小鼠視網膜無灌注區的相對面積較正常對照組明顯增大,差異有統計學意義(t=28.71,P<0.001)。
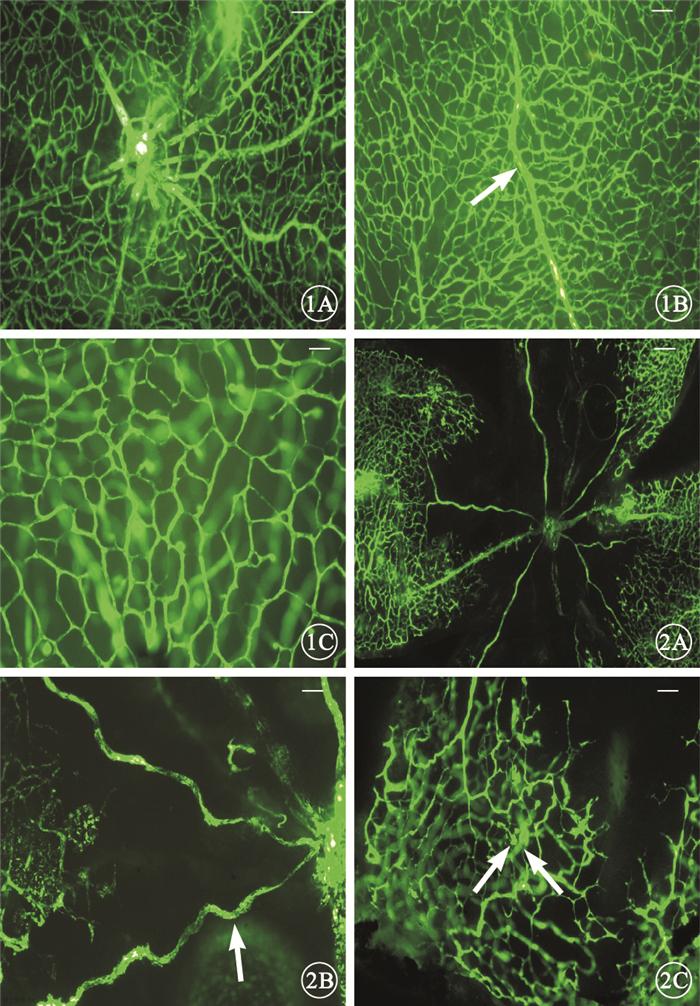 圖1
正常對照組小鼠視網膜熒光顯微鏡像。1A.視盤周圍血管分布及形態正常標尺:200μm;1B.自視盤發出的大血管正常走行(白箭)標尺:100μm;1C.視網膜血管分布呈均勻的網狀結構標尺:50μm ??圖 2 OIR組小鼠視網膜熒光顯微鏡像。2A.視盤周圍血管紆曲,分布紊亂標尺:200μm;2B.自視盤發出的大血管明顯紆曲(白箭)標尺:100μm;2C.可見大量的新生血管和新生血管芽(白箭),新生血管網結構及分布紊亂標尺:100μm
圖1
正常對照組小鼠視網膜熒光顯微鏡像。1A.視盤周圍血管分布及形態正常標尺:200μm;1B.自視盤發出的大血管正常走行(白箭)標尺:100μm;1C.視網膜血管分布呈均勻的網狀結構標尺:50μm ??圖 2 OIR組小鼠視網膜熒光顯微鏡像。2A.視盤周圍血管紆曲,分布紊亂標尺:200μm;2B.自視盤發出的大血管明顯紆曲(白箭)標尺:100μm;2C.可見大量的新生血管和新生血管芽(白箭),新生血管網結構及分布紊亂標尺:100μm
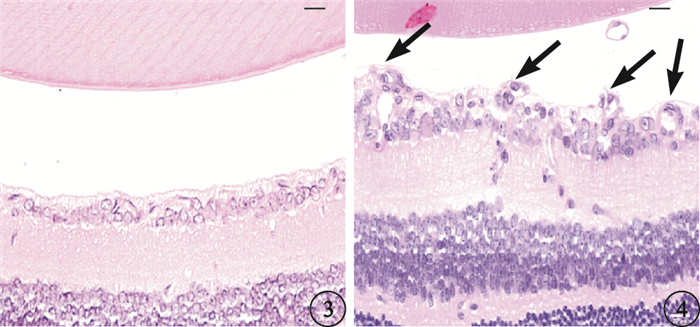
光學顯微鏡觀察發現,正常對照組小鼠內界膜結構完整、平滑,血管內皮細胞排列整齊,偶見突破內界膜的血管內皮細胞核(圖 3)。OIR組小鼠內界膜形態不規則,內界膜下細胞增生明顯,排列紊亂,可見大量突破內界膜呈簇狀排列的血管內皮細胞核組成新生血管管腔(圖 4)。正常對照組、OIR組平均每張切片突破內界膜的視網膜血管內皮細胞核數分別為(0.16±0.31)、(28.41±4.01)個。兩組平均每張切片突破內界膜的視網膜血管內皮細胞核數比較,差異有統計學意義(t=54.45,P<0.001)。
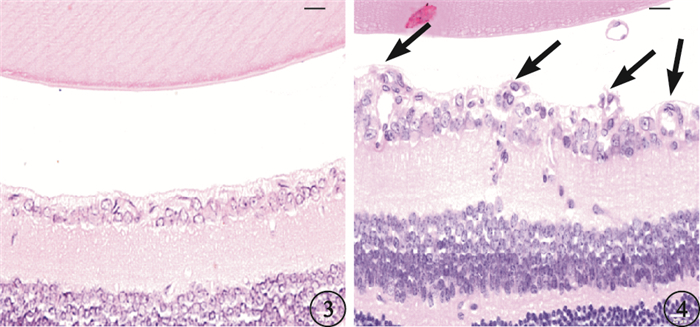 圖3
正常對照組小鼠視網膜光學顯微鏡像。未見突破內界膜的血管內皮細胞核HE標尺:20μm ??圖 4 OIR組小鼠視網膜光學顯微鏡像。可見大量突破內界膜的血管內皮細胞核(黑箭) HE標尺:20μm
圖3
正常對照組小鼠視網膜光學顯微鏡像。未見突破內界膜的血管內皮細胞核HE標尺:20μm ??圖 4 OIR組小鼠視網膜光學顯微鏡像。可見大量突破內界膜的血管內皮細胞核(黑箭) HE標尺:20μm
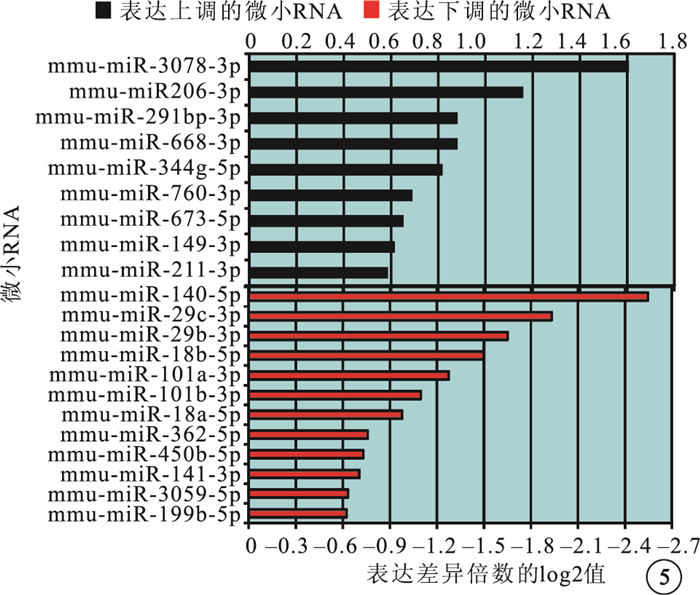
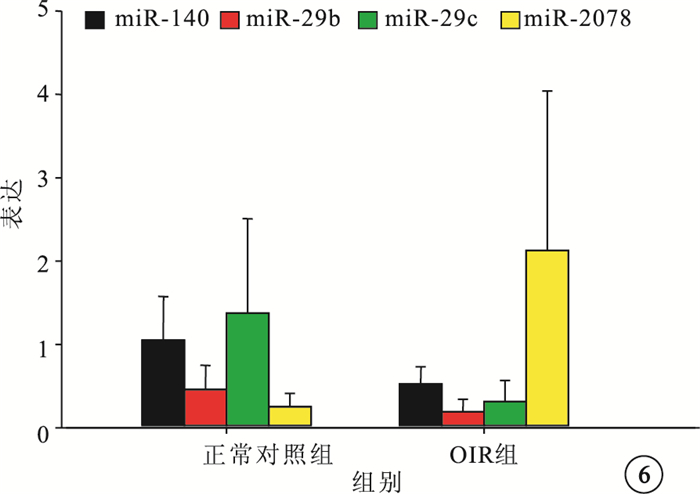
微小RNA芯片分析結果顯示,與正常對照組比較,OIR組有21個微小RNA的表達發生了1.5倍以上的變化。其中,上調者9個,下調者12個(圖 5)。差異表達倍數≥3.0的微小RNA為miR-3078、miR-140、miR-29b、miR-29c。RT-PCR檢測發現,差異表達倍數≥3.0的4個微小RNA的表達趨勢與芯片分析結果一致。與正常對照組比較,OIR組miR-3078的表達明顯上調,差異有統計學意義(t=-2.380,P<0.05);miR-140、miR-29b、miR-29c的表達明顯下調,差異也有統計學意義(t=2.638、2.323、2.415,P<0.05)(圖 6)。
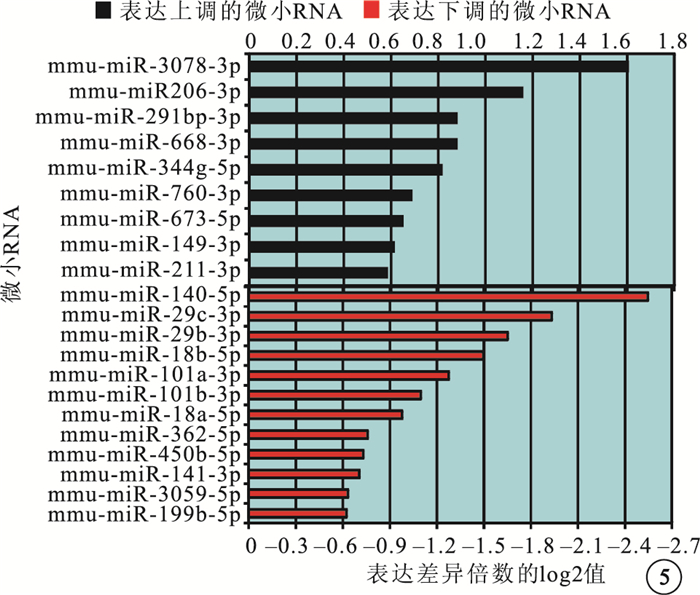 圖5
小鼠眼球微小RNA芯片分析結果
圖5
小鼠眼球微小RNA芯片分析結果
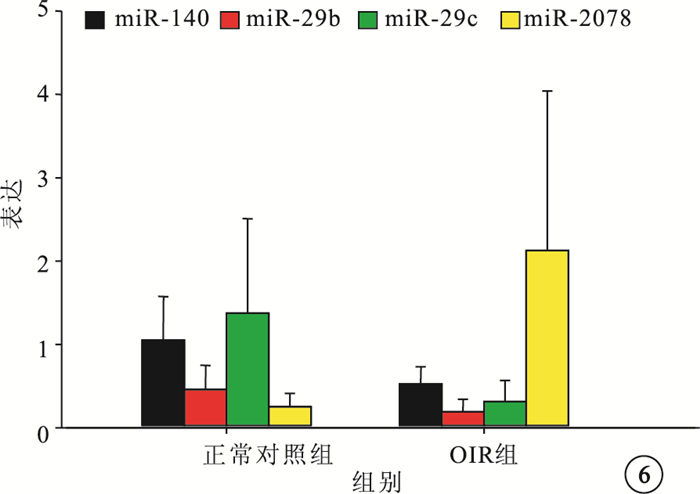 圖6
正常對照組和OIR組4個表達差異≥3.0倍以上的微小RNA表達比較。*與正常對照組比較,P<0.05
圖6
正常對照組和OIR組4個表達差異≥3.0倍以上的微小RNA表達比較。*與正常對照組比較,P<0.05
3 討論
OIR動物模型是研究RNV性眼病的主要模型之一[10]。視網膜中央有明顯的無灌注區、中央大血管走形紆曲、突破內界膜的血管細胞核明顯增多均是判斷OIR動物模型成功與否的重要指標[6, 11]。研究表明,出生后17 d是OIR小鼠RNV形成的高峰期[12]。我們參照Smith等[6]的方法建立了OIR小鼠模型,在小鼠17日齡時通過視網膜鋪片和石蠟切片HE染色發現,OIR組小鼠視網膜可見大面積無灌注區、新生血管及突破視網膜內界膜的血管內皮細胞核,證實模型制備成功。
近年來,微小RNA在眼部的表達和功能已逐漸被大家所認識[5]。研究發現,微小RNA可調節眼部組織的發育分化、損傷后再生、眼部細胞代謝和凋亡以及調節視網膜的生物節律等[13-18]。而有關微小RNA在RNV狀態下的表達也有報道。Shen等[19]在OIR小鼠模型中發現了7個表達上調的微小RNA(miR-106、-146、-181、-199a、-214、-424、-451)和3個表達下調的微小RNA(miR-31、-150、-184),并驗證了miR-31的可能靶基因是血小板來源的生長因子(PDGFB)和缺氧誘導因子1α(HIF-1α),miR-150的可能靶基因是VEGF和PDGFB,miR-184的可能靶基因是Wnt蛋白的受體分子Frizzled4。說明一種微小RNA可能調控多種蛋白質的翻譯過程,多個微小RNA也可能作用于同一種mRNA[20]。提示單個微小RNA的表達改變所導致的諸多基因的表達變化則可能為視網膜基因表達和功能調控提供有效的機制。我們利用微小RNA芯片技術比較了OIR小鼠和正常小鼠視網膜之間微小RNA的差異表達,發現了具有顯著表達差異的4個微小RNA,分別是miR-3078、miR-140、miR-29b、miR-29c。其中,miR-3078的表達顯著上調,miR-140、miR-29b和miR-29c的表達明顯下調。這些差異表達的微小RNA與眼部疾病及病理特征的關系將為RNV機制的深入研究提供思路。
miR-29b和miR-29c均屬于miR-29家族,具有相似的生物學行為,可調控參與代謝、分化、凋亡、遷移、免疫等過程的基因表達[21]。Loscher等[22]在視網膜色素變性鼠模型中發現,miR-29c在視網膜內核層的外層和光感受器中均有表達。Silva等[18]則在糖尿病視網膜病變鼠模型中發現。miRNA-29b在視網膜神經節細胞層及內核層中均有表達,而且還發現miR-29家族尤其是miR-29b的可能下游靶點蛋白激酶(PKR)信號通路相關蛋白X(RAX),并推測miR-29b可能通過參與PKR信號通路間接調控RAX蛋白,從而調控視網膜神經元細胞凋亡。Luna等[23]研究發現,miR-29b可負向調節小梁網細胞胞外基質的多種基因的表達,可能是慢性氧化應激條件下眼內壓升高的原因之一。說明miR-29家族可能與眼部氧化應激及炎癥有關,這恰好與RNV形成有關的缺血、缺氧及炎癥因素一致。提示miR-29家族可能直接或間接作用于與RNV的形成。目前有關MiR-140在眼部的作用鮮有報道,但其在抑制腫瘤生長和促進骨骼發育等方面有重要的調控作用[24]。研究發現,miR-140能夠靶向調控與新生血管形成關系密切的轉錄生長因子β受體1和成纖維細胞生長因子9基因的表達,從而抑制肝癌細胞的生長和遷移[25-27]。提示miR140可能通過調控血管生成因子的表達參與RNV形成,但具體機制有待進一步研究和證實。MiR-3078是本研究芯片檢測結果中具有明顯差異表達的唯一的上調微小RNA。由于其為較新發現的微小RNA,功能尚不清楚。我們推測本研究發現的4個差異表達的微小RNA可能參與了RNV形成,其機制可能與缺血缺氧、氧化應激及炎癥導致的血管生成因子表達增加或相應信號通路的激活有關。
通過對OIR小鼠視網膜的微小RNA芯片分析,本研究發現了可能與RNV形成相關的微小RNA,這為進一步研究以新生血管為主要表現的視網膜病變的調控機制和治療靶點提供了新的思路。下一步我們將預測各差異表達微小RNA的下游靶點并通過分子生物學的方法對所預測的靶點進行驗證,逐步探索RNV形成的相關分子機制。
玻璃體腔注射抗血管內皮生長因子(VEGF)藥物是目前治療眼部新生血管性疾病的有效方法,但其存在并發癥、副作用多等問題,甚至部分患者經抗VEGF藥物治療后仍不能阻止視網膜新生血管(RNV)的形成[1-3]。尋找不依賴VEGF的促新生血管生成的其他通路,以及高效、低損傷拮抗RNV的治療靶點尤為重要。微小RNA由具有發夾結構的單鏈RNA前體經Dicer酶加工后生成,主要通過對靶mRNA的降解或翻譯抑制,使特定靶基因沉默而發揮作用[4]。目前已明確微小RNA在眼部組織發育分化、細胞凋亡、代謝和損傷后再生等方面具有調控作用[5]。但其是否在RNV形成過程中發揮作用還不清楚。為此,我們通過建立氧誘導視網膜病變(OIR)小鼠模型,利用微小RNA芯片分析技術檢測出具有差異表達的微小RNA并進行實時聚合酶鏈反應(RT-PCR)驗證,以探索與RNV形成有關的微小RNA。現將結果報道如下。
1 材料和方法
7日齡健康清潔級C57BL/6J小鼠52只,雌雄不限,由中國人民解放軍軍事醫學科學院實驗動物中心提供。按隨機數字表法將小鼠分為正常對照組和OIR組,每組26只。參照文獻[6]的方法建立OIR動物模型,將OIR組小鼠與哺乳母鼠一同置入密閉玻璃容器中,氣體流量控制在2 L/min左右,箱內含氧體積分數為(75±2)%;用CY-12C數字測氧儀(浙江建德梅城電化分析儀器廠)間斷性監測出氣口處氧濃度,6~8次/d。每日打開密閉容器1次,更換墊料、加食水,替換母鼠。高氧環境下連續飼養5 d后,再置于正常空氣中飼養5 d。正常對照組小鼠在正常空氣中飼養10 d,不做任何處理。
小鼠17日齡時,每組選取8只小鼠作視網膜鋪片,觀察小鼠視網膜血管形態,測量視網膜無灌注區的相對面積。參照文獻[7]的方法,取25 mg相對分子質量為2×106的異硫氰酸熒光素-葡萄聚糖(FITC-dextran)粉末(美國Sigma公司)溶于0.5 ml去離子水中,以離心半徑5.13 cm,13 000 r/min離心5 min,取上清液,制備50 mg/ml的FITC-dextran灌注液。以左手拇指及中指捏住小鼠頭頸部皮膚,食指輕壓眼眶使眼球突出,然后將依據小鼠眼球大小彎成弧形的1 ml注射器針頭刺入左眼內側眼角的Tenon氏筋膜,沿鼻側眼眶壁平行地對喉頭方向推進約2 mm達球后靜脈叢,略轉動注射器并去除眼后部壓力,緩慢注入0.05 ml的FITC-dextran灌注液。靜置5 min,頸椎脫臼法處死小鼠,摘除眼球置于4%的多聚甲醛液中固定40 min,之后放入磷酸鹽緩沖液中4℃避光保存。手術顯微鏡下,沿角鞏膜緣剪開眼球,去除角膜、虹膜,保留晶狀體在眼杯中。兩把鑷子配合牽拉眼杯后極部殘留的視神經,使鞏膜和脈絡膜與視網膜完整分離。去除晶狀體,將分離的視網膜以視盤為中心,放射狀切開4片,玻璃體面向上平鋪于載玻片上,用含4′, 6-二脒基-2-苯基吲哚(DAPI)的抗淬滅封片劑(美國Vectashield公司)封片,蓋上蓋玻片。熒光顯微鏡(日本Olympus公司)下觀察并照相。采用CellSens Standard軟件(日本Olympus公司)測量視網膜無灌注區占全視網膜的相對面積。
小鼠17日齡時,每組選取8只小鼠作視網膜石蠟切片,蘇木精-伊紅(HE)染色并計數突破視網膜內界膜的視網膜血管細胞核數。取小鼠眼球,常規方法制作蠟塊,行矢狀位3μm厚連續切片,相鄰2張切片間隔40μm,每只眼球取10張代表性切片行HE染色,排除含視神經的切片。統計平均每只眼球每張切片中突破內界膜的視網膜血管細胞核數。參照文獻[8, 9]的計數標準,僅計數與視網膜有緊密聯系的血管細胞核,不包括玻璃體腔內與內界膜無聯系的血管細胞核。
小鼠17日齡時,每組選取10只小鼠處死并取眼球,立即放于液氮中,并轉存于-80℃冰箱中。其中5只小鼠的眼球標本由廣州復能基因有限公司對80個微小RNA行芯片分析。另外5只小鼠的眼球在取得微小RNA芯片結果后采用RT-PCR驗證芯片檢測差異表達倍數≥3.0的微小RNA。采用E.Z.N.A.微小RNA Kit提取試劑盒(美國Omega Bio-Tek公司)提取微小RNA。將眼球組織放入預冷的玻璃管中研磨,并加入1 ml裂解液,冰上勻漿3 min,移至1.5 ml離心管中,室溫下孵育3 min。1 ml裂解液中加入200μl氯仿,蓋好管蓋后劇烈震蕩混勻,冰中孵育10 min, 以12 000×g在4℃條件下離心15 min。取上清液,并加入無水乙醇混勻,經試劑盒中的HiBind RNA Mini柱過濾去除mRNA,濾液再經過MicroElute RNA柱,使微小RNA吸附于過濾柱的濾膜上,最后用30μl洗脫液洗脫。Nanodrop 2000分光光度計測定microRNAs的含量和純度,-80℃保存待用。All-in-One? miRNA qRT-PCR Detection Kit試劑盒(Cat.No.AOMD-Q050,美國GeneCopoeia公司)逆轉微小RNA并進行PCR反應。取500 ng微小RNA,按說明書中的反應體系和操作步驟進行逆轉。根據微小RNA的濃度計算加入微小RNA的體積,并向離心管中依次加入微小RNA、PolyA聚合酶、逆轉錄酶、5倍反應試劑,并用滅菌水補足至25μl,渦旋混勻并離心后使用GeneAmp PCR System 2400擴增儀進行逆轉錄。參數:37℃孵育60 min,85℃變性5 min。合成的cDNA于-80℃保存備用。cDNA經5倍稀釋后與上、下游引物一起進行RT-PCR反應,其中以RNU6為內參基因。引物均購自廣州復能基因有限公司,分別為miR-3078(Cat#MmiRQP1700)、miR-140(Cat#MmiRQP0181)、miR-29b(Cat#MmiRQP0373)、miR-29c(Cat#MmiRQP0375)和RNU6(Cat#MmiRQP9002)。RT-PCR采用7900HT熒光定量PCR儀。擴增參數:50℃孵育2 min,95℃變性10 min,然后40次循環;95℃變性15 s,60℃退火和延伸1 min;并加入解離階段:95℃變性15 s,60℃退火15 s,然后95℃反應15 s,以判定擴增產物的特異性。擴增后,獲得各基因擴增的循環閾值(Ct),以2-ΔΔCt法分析所得數據。ΔCt=目的基因Ct-內參基因Ct,ΔCt=OIR組目的基因2ΔCt-正常對照組目的基因ΔCt。MicroRNA表達差異倍數用2-ΔΔCt計算,并以表達差異倍數的log2值作圖。
采用SPSS 20.0統計軟件行統計學分析處理。各組量化的實驗數據資料經Shapiro-Wilk檢驗呈正態分布,以均數±標準差(
2 結果
熒光顯微鏡觀察發現,正常對照組小鼠自視盤發出的大血管向四周呈放射狀均勻分布,直至視網膜周邊部,視網膜血管分布呈均勻的網狀結構(圖 1)。OIR組小鼠自視盤發出的大血管明顯紆曲,分支減少;中央可見大片無灌注區,中周部可見明顯的視網膜血管無灌注區和灌注區的分界,分界處可見大量的新生血管和新生血管芽,新生血管網結構及分布紊亂(圖 2)。定量分析結果顯示,正常對照組、OIR組小鼠視網膜無灌注區的相對面積分別為(6.57±3.6)%、(25.81±2.12)%。OIR組小鼠視網膜無灌注區的相對面積較正常對照組明顯增大,差異有統計學意義(t=28.71,P<0.001)。
圖1
正常對照組小鼠視網膜熒光顯微鏡像。1A.視盤周圍血管分布及形態正常標尺:200μm;1B.自視盤發出的大血管正常走行(白箭)標尺:100μm;1C.視網膜血管分布呈均勻的網狀結構標尺:50μm ??圖 2 OIR組小鼠視網膜熒光顯微鏡像。2A.視盤周圍血管紆曲,分布紊亂標尺:200μm;2B.自視盤發出的大血管明顯紆曲(白箭)標尺:100μm;2C.可見大量的新生血管和新生血管芽(白箭),新生血管網結構及分布紊亂標尺:100μm
光學顯微鏡觀察發現,正常對照組小鼠內界膜結構完整、平滑,血管內皮細胞排列整齊,偶見突破內界膜的血管內皮細胞核(圖 3)。OIR組小鼠內界膜形態不規則,內界膜下細胞增生明顯,排列紊亂,可見大量突破內界膜呈簇狀排列的血管內皮細胞核組成新生血管管腔(圖 4)。正常對照組、OIR組平均每張切片突破內界膜的視網膜血管內皮細胞核數分別為(0.16±0.31)、(28.41±4.01)個。兩組平均每張切片突破內界膜的視網膜血管內皮細胞核數比較,差異有統計學意義(t=54.45,P<0.001)。
圖3
正常對照組小鼠視網膜光學顯微鏡像。未見突破內界膜的血管內皮細胞核HE標尺:20μm ??圖 4 OIR組小鼠視網膜光學顯微鏡像。可見大量突破內界膜的血管內皮細胞核(黑箭) HE標尺:20μm
微小RNA芯片分析結果顯示,與正常對照組比較,OIR組有21個微小RNA的表達發生了1.5倍以上的變化。其中,上調者9個,下調者12個(圖 5)。差異表達倍數≥3.0的微小RNA為miR-3078、miR-140、miR-29b、miR-29c。RT-PCR檢測發現,差異表達倍數≥3.0的4個微小RNA的表達趨勢與芯片分析結果一致。與正常對照組比較,OIR組miR-3078的表達明顯上調,差異有統計學意義(t=-2.380,P<0.05);miR-140、miR-29b、miR-29c的表達明顯下調,差異也有統計學意義(t=2.638、2.323、2.415,P<0.05)(圖 6)。
圖5
小鼠眼球微小RNA芯片分析結果
圖6
正常對照組和OIR組4個表達差異≥3.0倍以上的微小RNA表達比較。*與正常對照組比較,P<0.05
3 討論
OIR動物模型是研究RNV性眼病的主要模型之一[10]。視網膜中央有明顯的無灌注區、中央大血管走形紆曲、突破內界膜的血管細胞核明顯增多均是判斷OIR動物模型成功與否的重要指標[6, 11]。研究表明,出生后17 d是OIR小鼠RNV形成的高峰期[12]。我們參照Smith等[6]的方法建立了OIR小鼠模型,在小鼠17日齡時通過視網膜鋪片和石蠟切片HE染色發現,OIR組小鼠視網膜可見大面積無灌注區、新生血管及突破視網膜內界膜的血管內皮細胞核,證實模型制備成功。
近年來,微小RNA在眼部的表達和功能已逐漸被大家所認識[5]。研究發現,微小RNA可調節眼部組織的發育分化、損傷后再生、眼部細胞代謝和凋亡以及調節視網膜的生物節律等[13-18]。而有關微小RNA在RNV狀態下的表達也有報道。Shen等[19]在OIR小鼠模型中發現了7個表達上調的微小RNA(miR-106、-146、-181、-199a、-214、-424、-451)和3個表達下調的微小RNA(miR-31、-150、-184),并驗證了miR-31的可能靶基因是血小板來源的生長因子(PDGFB)和缺氧誘導因子1α(HIF-1α),miR-150的可能靶基因是VEGF和PDGFB,miR-184的可能靶基因是Wnt蛋白的受體分子Frizzled4。說明一種微小RNA可能調控多種蛋白質的翻譯過程,多個微小RNA也可能作用于同一種mRNA[20]。提示單個微小RNA的表達改變所導致的諸多基因的表達變化則可能為視網膜基因表達和功能調控提供有效的機制。我們利用微小RNA芯片技術比較了OIR小鼠和正常小鼠視網膜之間微小RNA的差異表達,發現了具有顯著表達差異的4個微小RNA,分別是miR-3078、miR-140、miR-29b、miR-29c。其中,miR-3078的表達顯著上調,miR-140、miR-29b和miR-29c的表達明顯下調。這些差異表達的微小RNA與眼部疾病及病理特征的關系將為RNV機制的深入研究提供思路。
miR-29b和miR-29c均屬于miR-29家族,具有相似的生物學行為,可調控參與代謝、分化、凋亡、遷移、免疫等過程的基因表達[21]。Loscher等[22]在視網膜色素變性鼠模型中發現,miR-29c在視網膜內核層的外層和光感受器中均有表達。Silva等[18]則在糖尿病視網膜病變鼠模型中發現。miRNA-29b在視網膜神經節細胞層及內核層中均有表達,而且還發現miR-29家族尤其是miR-29b的可能下游靶點蛋白激酶(PKR)信號通路相關蛋白X(RAX),并推測miR-29b可能通過參與PKR信號通路間接調控RAX蛋白,從而調控視網膜神經元細胞凋亡。Luna等[23]研究發現,miR-29b可負向調節小梁網細胞胞外基質的多種基因的表達,可能是慢性氧化應激條件下眼內壓升高的原因之一。說明miR-29家族可能與眼部氧化應激及炎癥有關,這恰好與RNV形成有關的缺血、缺氧及炎癥因素一致。提示miR-29家族可能直接或間接作用于與RNV的形成。目前有關MiR-140在眼部的作用鮮有報道,但其在抑制腫瘤生長和促進骨骼發育等方面有重要的調控作用[24]。研究發現,miR-140能夠靶向調控與新生血管形成關系密切的轉錄生長因子β受體1和成纖維細胞生長因子9基因的表達,從而抑制肝癌細胞的生長和遷移[25-27]。提示miR140可能通過調控血管生成因子的表達參與RNV形成,但具體機制有待進一步研究和證實。MiR-3078是本研究芯片檢測結果中具有明顯差異表達的唯一的上調微小RNA。由于其為較新發現的微小RNA,功能尚不清楚。我們推測本研究發現的4個差異表達的微小RNA可能參與了RNV形成,其機制可能與缺血缺氧、氧化應激及炎癥導致的血管生成因子表達增加或相應信號通路的激活有關。
通過對OIR小鼠視網膜的微小RNA芯片分析,本研究發現了可能與RNV形成相關的微小RNA,這為進一步研究以新生血管為主要表現的視網膜病變的調控機制和治療靶點提供了新的思路。下一步我們將預測各差異表達微小RNA的下游靶點并通過分子生物學的方法對所預測的靶點進行驗證,逐步探索RNV形成的相關分子機制。