引用本文: 馬維, 汪曉磊, 章欣欣, 陳璐勔, 韓松, 李俊發. 轉化生長因子-β在激光誘導脈絡膜新生血管形成中的作用. 中華眼底病雜志, 2014, 30(6): 594-598. doi: 10.3760/cma.j.issn.1005-1015.2014.06.014 復制
脈絡膜新生血管(CNV)因其累及視網膜黃斑區并引起反復出血、滲出和瘢痕形成,從而導致患者中心視力嚴重下降[1]。目前認為CNV形成的主要機制可能與血管生成因子與抑制因子的失衡有關[2]。作為血管生成因子之一的轉化生長因子(TGF)-β,與組織纖維化和瘢痕形成之間的關系最為密切,同時對炎性細胞和成纖維細胞有較強的趨化作用。TGF-β可通過調節血管內皮細胞的活性和血管內皮生長因子(VEGF)的表達,從而促進新生血管的發生[3]。但目前有關TGF-β在CNV形成過程中的具體作用及其機制尚不完全明確。為此,我們通過激光誘導小鼠脈絡膜CNV形成,探討TGF-β在CNV形成中的作用及其與VEGF和腫瘤壞死因子(TNF)-α的關系。現將結果報道如下。
1 材料和方法
6~8周齡雄性C57BL/6J小鼠80只,體重20~30 g,SPF級,中國醫學科學院實驗動物所提供。實驗動物飼養于安靜清潔環境,保證食物水源充足,環境相對濕度50%~60%,溫度18℃~22℃。TGF-β受體抑制劑LY2157299(美國Selleck公司),鼠源性TGF-β單克隆抗體(美國R&D公司),兔源性VEGF和TNF-α多克隆抗體(美國Abcam公司),鼠源性β-肌動蛋白(actin)單克隆抗體(美國Proteintech公司),異硫氰酸熒光素-同工凝集素B4 (FITC-isolectin B4,美國Vector Laboratories公司),辣根過氧化物酶(HRP)標記山羊抗兔IgG、HRP標記山羊抗鼠IgG(北京中杉金橋生物技術有限公司),紅霉素眼膏(上海通用藥業公司),歷設得熒光素鈉注射液(美國Alcon公司),5%托吡卡胺眼液(日本參天制藥株式會社),0.5%愛爾凱因眼液(英國Alcon公司),卡波姆滴眼液(美國Bausch & Lomb公司)。
實驗操作均遵循美國視覺與眼科學會關于眼科和視覺科學實驗動物使用規范,并取得首都醫科大學動物管理委員會的許可。將小鼠隨機分為正常對照組、光凝模型組、光凝加磷酸鹽緩沖液(PBS)對照組(PBS對照組)、光凝加TGF-β受體抑制劑組(TGF-β受體抑制劑組),每組20只。光凝模型組、PBS對照組及TGF-β受體抑制劑組小鼠均應用激光光凝誘導CNV形成,建立動物模型。實驗前首先排除動物眼部或全身性病變。激光參數:直徑50 μm、曝光時間0.1 s、功率150 mW(Novus Spectra高功率532激光系統,美國Lumenis公司)。小鼠麻醉后圍繞其視盤周圍在血管間光凝,以氣泡產生提示Bruch膜被擊中為準[4]。建模后,依據藥物說明書的方法,TGF-β受體抑制劑組小鼠按體重3.0 μg/g的藥物劑量腹腔注射LY2157299,1次/d;PBS對照組注射同等劑量的含有0.06%二甲基亞砜的PBS。激光光凝后1、2、3、4周,正常對照組及光凝模型組小鼠腹腔注射5%水合氯醛充分麻醉,復方托吡卡胺滴眼液充分散大瞳孔,腹腔注射2%熒光素鈉0.1 ml,應用眼底血管造影機(德國Heidelberg公司)行熒光素眼底血管造影(FFA)檢查,同步動態觀察小鼠模型CNV的形成情況。
激光光凝后1、2、3、4周,采用蛋白免疫印跡法(Western blot)檢測正常對照組及光凝模型組小鼠視網膜內TGF-β的蛋白表達以及正常對照組、PBS對照組、TGF-β受體抑制劑組小鼠視網膜內VEGF和TNF-α的蛋白表達。各組隨機選取小鼠5只,摘取眼球,完整分離視網膜組織,存放于預冷Eppendorf管并迅速移至液氮,凍存備用;應用美國Sigma公司提供的蛋白抽提試劑提取蛋白,用二喹啉甲酸法進行蛋白定量,配制樣品。取50 μg總蛋白樣品,進行十二烷基硫酸鈉-聚丙烯酰胺凝膠蛋白電泳,然后將蛋白轉至聚偏氟乙烯(PVDF)膜上。10%脫脂牛奶封閉PVDF膜1 h,含0.05%吐溫20的三羥甲基氨基甲烷鹽酸緩沖鹽溶液(TTBS)漂洗3次,每次10 min。分別應用鼠源性TGF-β單克隆抗體(1 :1000)、兔源性VEGF多克隆抗體(1 :1000)室溫孵育3 h;兔源性TNF-α多克隆抗體(1 :1000) 4℃過夜,室溫孵育1 h;鼠源性β-actin單克隆抗體(1 :20 000)室溫孵育1 h。TTBS漂洗3次,每次10 min。應用HRP標記的二抗(1 :4000)室溫孵育1 h,TTBS漂洗3次,每次10 min。電化學發光法顯色發光,暗室內行X線片曝光。應用圖像分析軟件Quantity One對膠片中蛋白條帶進行灰度值分析,并以β-actin蛋白為內參,計算各組蛋白與β-actin的灰度比值,進行半定量分析。
激光光凝后2周,作視網膜色素上皮細胞(RPE)/脈絡膜組織鋪片,采用熒光染色計算CNV面積。各組隨機選取小鼠5只,摘取眼球,4%多聚甲醛固定1 h,沖洗后放入PBS溶液,解剖顯微鏡下去除眼前節并剝離視網膜。將RPE-脈絡膜-鞏膜復合體置于載玻片上,以視盤為中心對稱行4條放射狀切口。0.5%熒光Isolectin B4避光染色4℃過夜,PBS漂洗3次。鋪片、封片,用激光共聚焦顯微鏡對CNV進行觀察,測量面積。
采用SPSS 11.5統計軟件行統計學分析,所得數據以均數±標準差(
2 結果
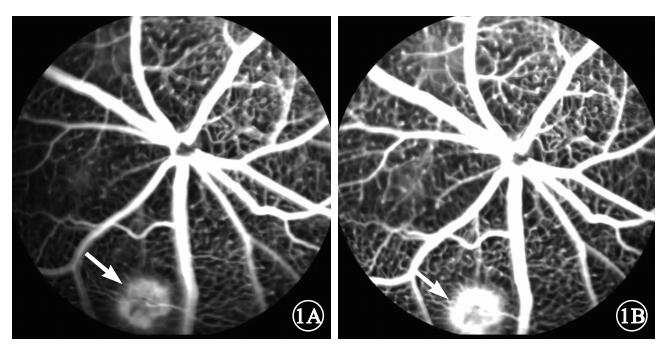
FFA檢查結果顯示,激光光凝后1周,光凝模型組小鼠動脈早期激光光凝區出現細網狀強熒光(圖 1A),造影過程中熒光強度增強,范圍擴大,部分強熒光片融合,晚期熒光素滲漏呈盤狀強熒光團(圖 1B)。
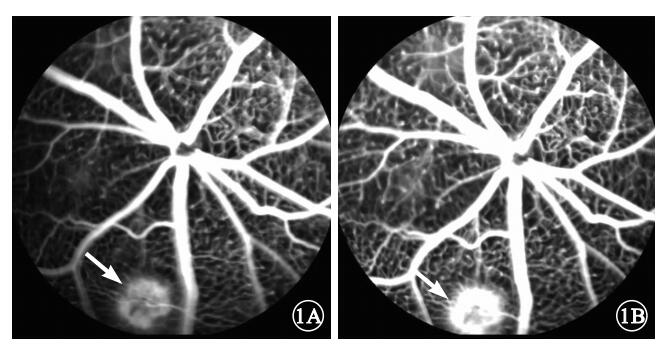 圖1
光凝模型組小鼠FFA像。1A.光凝區早期熒光素滲漏呈細網狀強熒光(白箭);1B.光凝區晚期部分熒光片融合呈盤狀強熒光團(白箭)
圖1
光凝模型組小鼠FFA像。1A.光凝區早期熒光素滲漏呈細網狀強熒光(白箭);1B.光凝區晚期部分熒光片融合呈盤狀強熒光團(白箭)
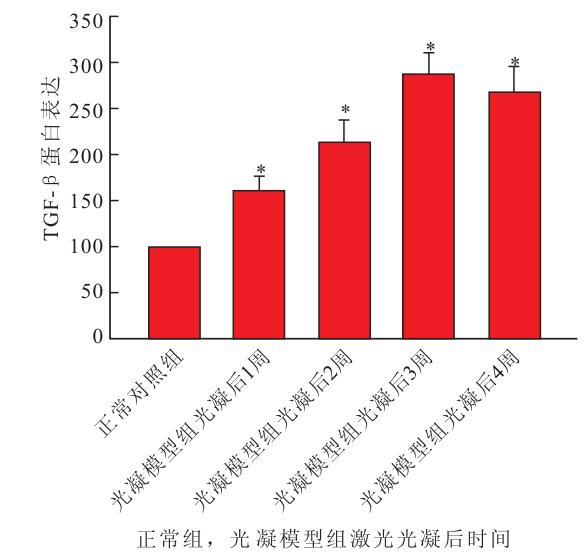
Western blot檢測結果顯示,正常對照組小鼠視網膜內可見相對分子質量為45×103的TGF-β蛋白表達;激光光凝后1周,光凝模型組小鼠視網膜內TGF-β蛋白表達開始升高,2~4周TGF-β蛋白表達增加明顯。隨著激光光凝的時間延長,光凝模型組小鼠視網膜內TGF-β蛋白表達逐漸升高,于激光光凝后3周達高峰,激光光凝后4周仍維持在高水平。光凝模型組與正常對照組小鼠視網膜內TGF-β蛋白表達比較,差異有統計學意義(F=13.042,P<0.05)(圖 2)。
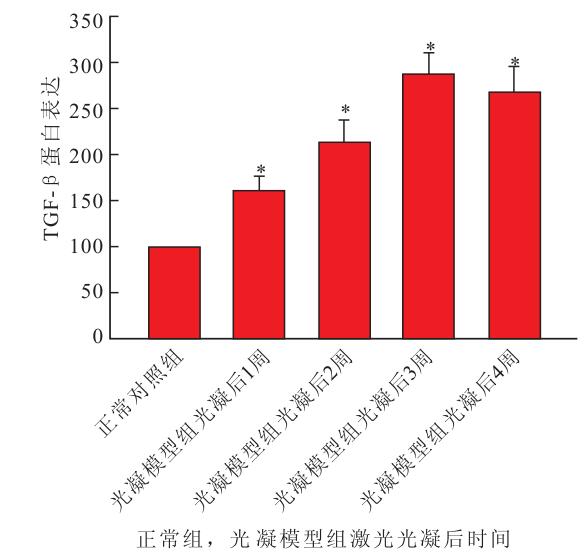 圖2
正常對照組與光凝模型組小鼠視網膜內TGF-β蛋白表達比較。*與正常對照組比較,P<0.05
圖2
正常對照組與光凝模型組小鼠視網膜內TGF-β蛋白表達比較。*與正常對照組比較,P<0.05
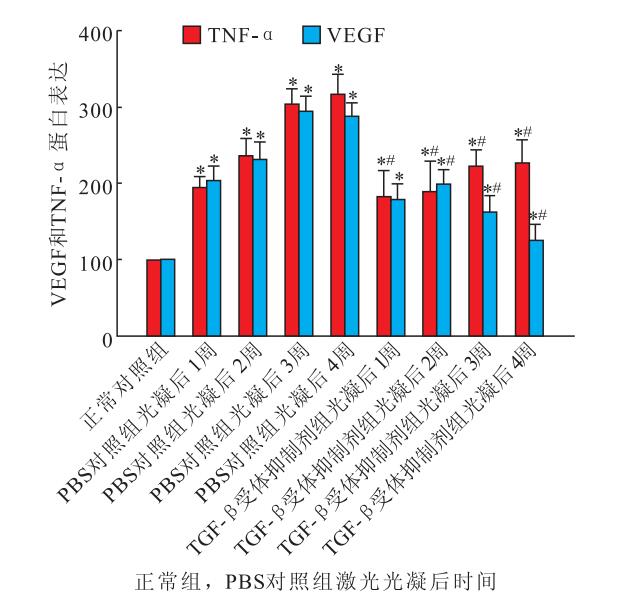
激光光凝后1、2、3、4周,PBS對照組、TGF-β受體抑制劑組小鼠視網膜內TNF-α(F=14.721、17.509)、VEGF(F=18.890、11.251)蛋白表達均較正常對照組明顯增加,差異均有統計學意義(P<0.05)。與PBS對照組比較,TGF-β受體抑制劑組小鼠視網膜內TNF-α、VEGF蛋白表達明顯降低,差異均有統計學意義(F=21.321、16.160,P<0.05)(圖 3)。
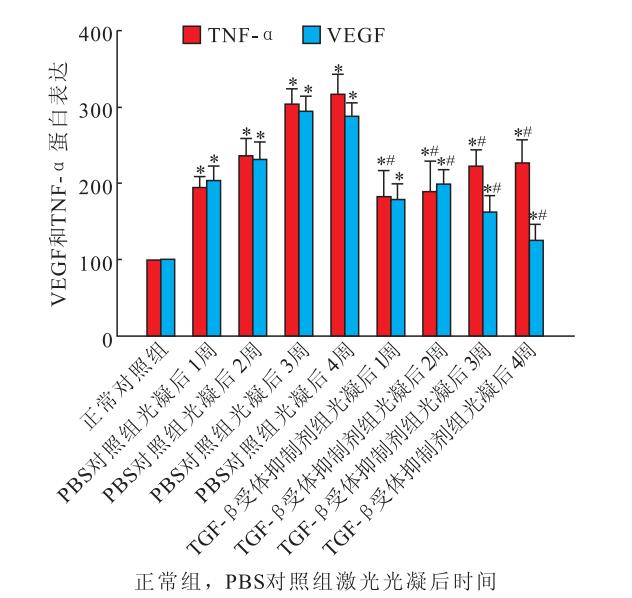 圖3
正常對照組、PBS對照組與TGF-β受體抑制劑組小鼠視網膜內TNF-α和VEGF蛋白表達比較。*與正常對照組比較,P<0.05;#與PBS對照組比較,P<0.05
圖3
正常對照組、PBS對照組與TGF-β受體抑制劑組小鼠視網膜內TNF-α和VEGF蛋白表達比較。*與正常對照組比較,P<0.05;#與PBS對照組比較,P<0.05
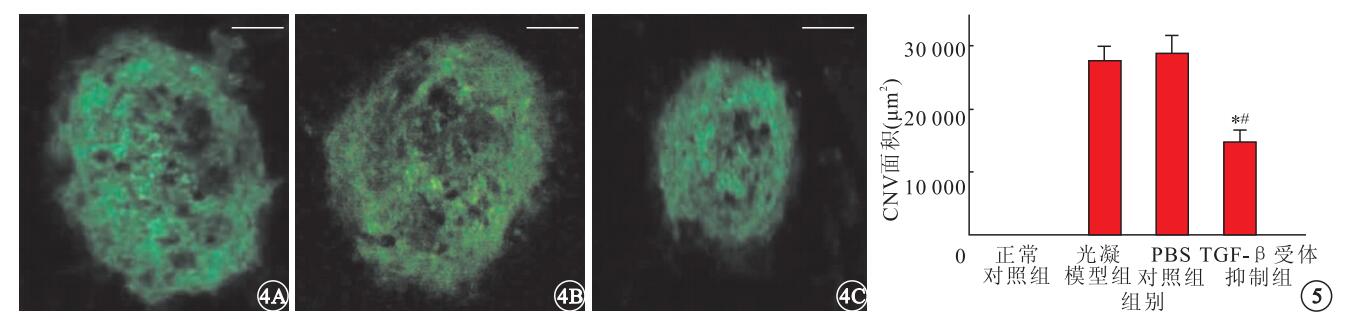
激光光凝后2周,熒光顯微鏡觀察發現,正常對照組小鼠脈絡膜內無新生血管形成;光凝模型組、PBS對照組及TGF-β受體抑制劑組小鼠脈絡膜內可見綠色強熒光團(圖 4)。光凝模型組、PBS對照組及TGF-β受體抑制劑組小鼠脈絡膜內CNV面積分別為(27 518.83±2 466.56)、(28 780.81±2 958.37)、(14 615.65±2 079.58) μm2;TGF-β受體抑制劑組小鼠脈絡膜內CNV面積明顯小于PBS對照組(F=4.482)及光凝模型組(F=11.374),差異均有統計學意義(P<0.05)(圖 5)。
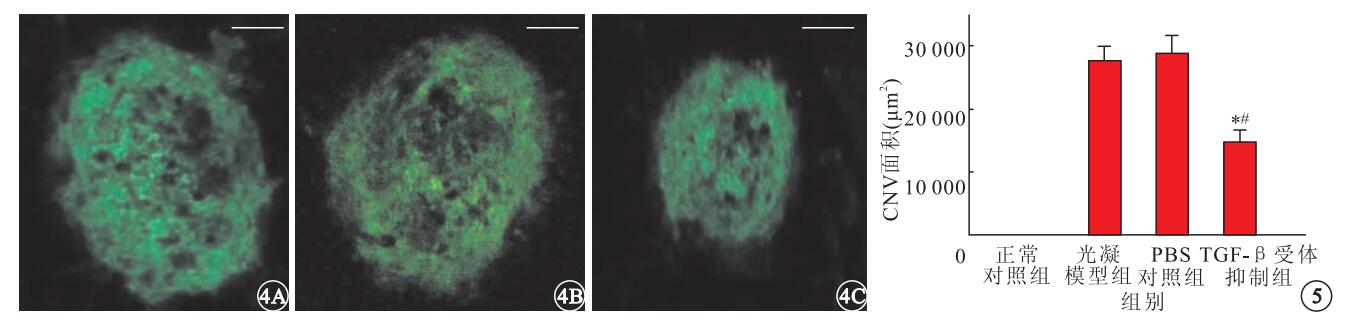 圖4
小鼠脈絡膜鋪片熒光顯微鏡像。4A.光凝模型組;4B. PBS對照組;4C. TGF-β受體抑制劑組。光凝模型組、PBS對照組及TGF-β受體抑制劑組小鼠脈絡膜內可見綠色強亮熒光團標尺:50 μm??圖 5??各組小鼠CNV面積比較。*與正常對照組比較,P<0.05;#與PBS對照組比較,P<0.05
圖4
小鼠脈絡膜鋪片熒光顯微鏡像。4A.光凝模型組;4B. PBS對照組;4C. TGF-β受體抑制劑組。光凝模型組、PBS對照組及TGF-β受體抑制劑組小鼠脈絡膜內可見綠色強亮熒光團標尺:50 μm??圖 5??各組小鼠CNV面積比較。*與正常對照組比較,P<0.05;#與PBS對照組比較,P<0.05
3 討論
激光誘導CNV形成建立動物模型成模率高、重復性好、成模時間短、成本低,易從分子水平研究CNV發病機制,對評價新型新生血管抑制劑的療效具有重要意義[5]。我們采用這一方法建立動物模型,發現激光光凝后1周小鼠脈絡膜內熒光素滲漏呈盤狀強熒光團。說明激光誘導CNV形成,小鼠模型建立成功。TGF-β生物學功能復雜,參與了細胞增生、分化與細胞外基質的代謝調節,具有抑制和刺激的雙重作用[6]。在體外,TGF-β對于成纖維細胞生長因子誘導的內皮細胞增生和趨化有抑制作用;在體內,TGF-β是一種血管生成因子,但當其濃度增高達到一定程度時,可抑制新生血管的形成。研究表明,在激光誘導的成年大鼠CNV模型新生血管內皮細胞內TGF-β mRNA轉錄水平增加[7];在老年性黃斑變性患眼的RPE中以及在手術切除的CNV膜轉分化為RPE細胞中,TGF-β表達增加[8]。本研究進一步觀察TGF-β在激光誘導小鼠CNV模型視網膜內表達情況,結果顯示激光光凝后小鼠視網膜內TGF-β蛋白表達量隨光凝時間延長而逐漸升高,并于光凝后3周達高峰。我們還發現,激光光凝后PBS對照組、TGF-β受體抑制劑組小鼠視網膜內TNF-α和VEGF蛋白表達均較正常對照組增高,而TGF-β受體抑制劑組小鼠視網膜內TNF-α和VEGF蛋白表達又較PBS對照組明顯降低。提示小鼠視網膜內TNF-α和VEGF蛋白表達增高可能是由于TGF-β高表達引起。
TGF-β的細胞漿內信號傳導通路主要包括膜受體絲氨酸/蘇氨酸激酶系統和Smad蛋白信號傳遞系統。TGF-β通過與3種高親和力的細胞膜表面受體結合,發揮其效應[9]。LY2157299是一種TGF-β受體激酶小分子抑制劑,主要用于腫瘤治療,其研究目前尚處于臨床前期階段。體外研究結果顯示,TGF-β I型受體激酶小分子抑制劑可以有效阻斷Smad磷酸化,限制腫瘤生長,減少腫瘤的轉移和侵襲[10]。本研究在小鼠CNV模型中應用該抑制劑,定量計算其對CNV生成面積的影響。此前有文獻報道,應用TGF-β抑制肽可以抑制激光誘導大鼠CNV的早期形成[11, 12]。但關于TGF-β抑制劑對激光誘導小鼠CNV及其視網膜內TNF-α和VEGF蛋白水平影響的研究還少見報道。本研究結果顯示,TGF-β受體抑制劑組小鼠視網膜內TNF-α和VEGF蛋白表達受到明顯抑制,且CNV面積明顯小于PBS對照組。表明TGF-β在CNV發生發展中發揮了促進作用,應用其抑制劑可以減少CNV形成。這可能是由于受體激酶信號傳遞的干擾阻礙了TGF-β的信號傳導,抑制其在新生血管形成中發揮正常生物學功能,在一定程度上抑制了細胞分泌VEGF[13],減少巨噬細胞在損傷部位的聚集,從而降低TNF-α等促血管生成因子的分泌,起到抑制CNV形成的作用。
本研究結果表明,通過調控TGF-β蛋白表達水平或干預其信號傳導通路,可能會成為抑制CNV形成的一個新的重要手段,從而為更加高效、長效的多因子聯合防治CNV提供理論和實驗依據。然而,TGF-β/Smad信號傳導通路在機體組織中作用廣泛,其與其他信號通路相互作用,抑制該信號通路對機體的影響有待進一步深入研究。
脈絡膜新生血管(CNV)因其累及視網膜黃斑區并引起反復出血、滲出和瘢痕形成,從而導致患者中心視力嚴重下降[1]。目前認為CNV形成的主要機制可能與血管生成因子與抑制因子的失衡有關[2]。作為血管生成因子之一的轉化生長因子(TGF)-β,與組織纖維化和瘢痕形成之間的關系最為密切,同時對炎性細胞和成纖維細胞有較強的趨化作用。TGF-β可通過調節血管內皮細胞的活性和血管內皮生長因子(VEGF)的表達,從而促進新生血管的發生[3]。但目前有關TGF-β在CNV形成過程中的具體作用及其機制尚不完全明確。為此,我們通過激光誘導小鼠脈絡膜CNV形成,探討TGF-β在CNV形成中的作用及其與VEGF和腫瘤壞死因子(TNF)-α的關系。現將結果報道如下。
1 材料和方法
6~8周齡雄性C57BL/6J小鼠80只,體重20~30 g,SPF級,中國醫學科學院實驗動物所提供。實驗動物飼養于安靜清潔環境,保證食物水源充足,環境相對濕度50%~60%,溫度18℃~22℃。TGF-β受體抑制劑LY2157299(美國Selleck公司),鼠源性TGF-β單克隆抗體(美國R&D公司),兔源性VEGF和TNF-α多克隆抗體(美國Abcam公司),鼠源性β-肌動蛋白(actin)單克隆抗體(美國Proteintech公司),異硫氰酸熒光素-同工凝集素B4 (FITC-isolectin B4,美國Vector Laboratories公司),辣根過氧化物酶(HRP)標記山羊抗兔IgG、HRP標記山羊抗鼠IgG(北京中杉金橋生物技術有限公司),紅霉素眼膏(上海通用藥業公司),歷設得熒光素鈉注射液(美國Alcon公司),5%托吡卡胺眼液(日本參天制藥株式會社),0.5%愛爾凱因眼液(英國Alcon公司),卡波姆滴眼液(美國Bausch & Lomb公司)。
實驗操作均遵循美國視覺與眼科學會關于眼科和視覺科學實驗動物使用規范,并取得首都醫科大學動物管理委員會的許可。將小鼠隨機分為正常對照組、光凝模型組、光凝加磷酸鹽緩沖液(PBS)對照組(PBS對照組)、光凝加TGF-β受體抑制劑組(TGF-β受體抑制劑組),每組20只。光凝模型組、PBS對照組及TGF-β受體抑制劑組小鼠均應用激光光凝誘導CNV形成,建立動物模型。實驗前首先排除動物眼部或全身性病變。激光參數:直徑50 μm、曝光時間0.1 s、功率150 mW(Novus Spectra高功率532激光系統,美國Lumenis公司)。小鼠麻醉后圍繞其視盤周圍在血管間光凝,以氣泡產生提示Bruch膜被擊中為準[4]。建模后,依據藥物說明書的方法,TGF-β受體抑制劑組小鼠按體重3.0 μg/g的藥物劑量腹腔注射LY2157299,1次/d;PBS對照組注射同等劑量的含有0.06%二甲基亞砜的PBS。激光光凝后1、2、3、4周,正常對照組及光凝模型組小鼠腹腔注射5%水合氯醛充分麻醉,復方托吡卡胺滴眼液充分散大瞳孔,腹腔注射2%熒光素鈉0.1 ml,應用眼底血管造影機(德國Heidelberg公司)行熒光素眼底血管造影(FFA)檢查,同步動態觀察小鼠模型CNV的形成情況。
激光光凝后1、2、3、4周,采用蛋白免疫印跡法(Western blot)檢測正常對照組及光凝模型組小鼠視網膜內TGF-β的蛋白表達以及正常對照組、PBS對照組、TGF-β受體抑制劑組小鼠視網膜內VEGF和TNF-α的蛋白表達。各組隨機選取小鼠5只,摘取眼球,完整分離視網膜組織,存放于預冷Eppendorf管并迅速移至液氮,凍存備用;應用美國Sigma公司提供的蛋白抽提試劑提取蛋白,用二喹啉甲酸法進行蛋白定量,配制樣品。取50 μg總蛋白樣品,進行十二烷基硫酸鈉-聚丙烯酰胺凝膠蛋白電泳,然后將蛋白轉至聚偏氟乙烯(PVDF)膜上。10%脫脂牛奶封閉PVDF膜1 h,含0.05%吐溫20的三羥甲基氨基甲烷鹽酸緩沖鹽溶液(TTBS)漂洗3次,每次10 min。分別應用鼠源性TGF-β單克隆抗體(1 :1000)、兔源性VEGF多克隆抗體(1 :1000)室溫孵育3 h;兔源性TNF-α多克隆抗體(1 :1000) 4℃過夜,室溫孵育1 h;鼠源性β-actin單克隆抗體(1 :20 000)室溫孵育1 h。TTBS漂洗3次,每次10 min。應用HRP標記的二抗(1 :4000)室溫孵育1 h,TTBS漂洗3次,每次10 min。電化學發光法顯色發光,暗室內行X線片曝光。應用圖像分析軟件Quantity One對膠片中蛋白條帶進行灰度值分析,并以β-actin蛋白為內參,計算各組蛋白與β-actin的灰度比值,進行半定量分析。
激光光凝后2周,作視網膜色素上皮細胞(RPE)/脈絡膜組織鋪片,采用熒光染色計算CNV面積。各組隨機選取小鼠5只,摘取眼球,4%多聚甲醛固定1 h,沖洗后放入PBS溶液,解剖顯微鏡下去除眼前節并剝離視網膜。將RPE-脈絡膜-鞏膜復合體置于載玻片上,以視盤為中心對稱行4條放射狀切口。0.5%熒光Isolectin B4避光染色4℃過夜,PBS漂洗3次。鋪片、封片,用激光共聚焦顯微鏡對CNV進行觀察,測量面積。
采用SPSS 11.5統計軟件行統計學分析,所得數據以均數±標準差(
2 結果
FFA檢查結果顯示,激光光凝后1周,光凝模型組小鼠動脈早期激光光凝區出現細網狀強熒光(圖 1A),造影過程中熒光強度增強,范圍擴大,部分強熒光片融合,晚期熒光素滲漏呈盤狀強熒光團(圖 1B)。
圖1
光凝模型組小鼠FFA像。1A.光凝區早期熒光素滲漏呈細網狀強熒光(白箭);1B.光凝區晚期部分熒光片融合呈盤狀強熒光團(白箭)
Western blot檢測結果顯示,正常對照組小鼠視網膜內可見相對分子質量為45×103的TGF-β蛋白表達;激光光凝后1周,光凝模型組小鼠視網膜內TGF-β蛋白表達開始升高,2~4周TGF-β蛋白表達增加明顯。隨著激光光凝的時間延長,光凝模型組小鼠視網膜內TGF-β蛋白表達逐漸升高,于激光光凝后3周達高峰,激光光凝后4周仍維持在高水平。光凝模型組與正常對照組小鼠視網膜內TGF-β蛋白表達比較,差異有統計學意義(F=13.042,P<0.05)(圖 2)。
圖2
正常對照組與光凝模型組小鼠視網膜內TGF-β蛋白表達比較。*與正常對照組比較,P<0.05
激光光凝后1、2、3、4周,PBS對照組、TGF-β受體抑制劑組小鼠視網膜內TNF-α(F=14.721、17.509)、VEGF(F=18.890、11.251)蛋白表達均較正常對照組明顯增加,差異均有統計學意義(P<0.05)。與PBS對照組比較,TGF-β受體抑制劑組小鼠視網膜內TNF-α、VEGF蛋白表達明顯降低,差異均有統計學意義(F=21.321、16.160,P<0.05)(圖 3)。
圖3
正常對照組、PBS對照組與TGF-β受體抑制劑組小鼠視網膜內TNF-α和VEGF蛋白表達比較。*與正常對照組比較,P<0.05;#與PBS對照組比較,P<0.05
激光光凝后2周,熒光顯微鏡觀察發現,正常對照組小鼠脈絡膜內無新生血管形成;光凝模型組、PBS對照組及TGF-β受體抑制劑組小鼠脈絡膜內可見綠色強熒光團(圖 4)。光凝模型組、PBS對照組及TGF-β受體抑制劑組小鼠脈絡膜內CNV面積分別為(27 518.83±2 466.56)、(28 780.81±2 958.37)、(14 615.65±2 079.58) μm2;TGF-β受體抑制劑組小鼠脈絡膜內CNV面積明顯小于PBS對照組(F=4.482)及光凝模型組(F=11.374),差異均有統計學意義(P<0.05)(圖 5)。
圖4
小鼠脈絡膜鋪片熒光顯微鏡像。4A.光凝模型組;4B. PBS對照組;4C. TGF-β受體抑制劑組。光凝模型組、PBS對照組及TGF-β受體抑制劑組小鼠脈絡膜內可見綠色強亮熒光團標尺:50 μm??圖 5??各組小鼠CNV面積比較。*與正常對照組比較,P<0.05;#與PBS對照組比較,P<0.05
3 討論
激光誘導CNV形成建立動物模型成模率高、重復性好、成模時間短、成本低,易從分子水平研究CNV發病機制,對評價新型新生血管抑制劑的療效具有重要意義[5]。我們采用這一方法建立動物模型,發現激光光凝后1周小鼠脈絡膜內熒光素滲漏呈盤狀強熒光團。說明激光誘導CNV形成,小鼠模型建立成功。TGF-β生物學功能復雜,參與了細胞增生、分化與細胞外基質的代謝調節,具有抑制和刺激的雙重作用[6]。在體外,TGF-β對于成纖維細胞生長因子誘導的內皮細胞增生和趨化有抑制作用;在體內,TGF-β是一種血管生成因子,但當其濃度增高達到一定程度時,可抑制新生血管的形成。研究表明,在激光誘導的成年大鼠CNV模型新生血管內皮細胞內TGF-β mRNA轉錄水平增加[7];在老年性黃斑變性患眼的RPE中以及在手術切除的CNV膜轉分化為RPE細胞中,TGF-β表達增加[8]。本研究進一步觀察TGF-β在激光誘導小鼠CNV模型視網膜內表達情況,結果顯示激光光凝后小鼠視網膜內TGF-β蛋白表達量隨光凝時間延長而逐漸升高,并于光凝后3周達高峰。我們還發現,激光光凝后PBS對照組、TGF-β受體抑制劑組小鼠視網膜內TNF-α和VEGF蛋白表達均較正常對照組增高,而TGF-β受體抑制劑組小鼠視網膜內TNF-α和VEGF蛋白表達又較PBS對照組明顯降低。提示小鼠視網膜內TNF-α和VEGF蛋白表達增高可能是由于TGF-β高表達引起。
TGF-β的細胞漿內信號傳導通路主要包括膜受體絲氨酸/蘇氨酸激酶系統和Smad蛋白信號傳遞系統。TGF-β通過與3種高親和力的細胞膜表面受體結合,發揮其效應[9]。LY2157299是一種TGF-β受體激酶小分子抑制劑,主要用于腫瘤治療,其研究目前尚處于臨床前期階段。體外研究結果顯示,TGF-β I型受體激酶小分子抑制劑可以有效阻斷Smad磷酸化,限制腫瘤生長,減少腫瘤的轉移和侵襲[10]。本研究在小鼠CNV模型中應用該抑制劑,定量計算其對CNV生成面積的影響。此前有文獻報道,應用TGF-β抑制肽可以抑制激光誘導大鼠CNV的早期形成[11, 12]。但關于TGF-β抑制劑對激光誘導小鼠CNV及其視網膜內TNF-α和VEGF蛋白水平影響的研究還少見報道。本研究結果顯示,TGF-β受體抑制劑組小鼠視網膜內TNF-α和VEGF蛋白表達受到明顯抑制,且CNV面積明顯小于PBS對照組。表明TGF-β在CNV發生發展中發揮了促進作用,應用其抑制劑可以減少CNV形成。這可能是由于受體激酶信號傳遞的干擾阻礙了TGF-β的信號傳導,抑制其在新生血管形成中發揮正常生物學功能,在一定程度上抑制了細胞分泌VEGF[13],減少巨噬細胞在損傷部位的聚集,從而降低TNF-α等促血管生成因子的分泌,起到抑制CNV形成的作用。
本研究結果表明,通過調控TGF-β蛋白表達水平或干預其信號傳導通路,可能會成為抑制CNV形成的一個新的重要手段,從而為更加高效、長效的多因子聯合防治CNV提供理論和實驗依據。然而,TGF-β/Smad信號傳導通路在機體組織中作用廣泛,其與其他信號通路相互作用,抑制該信號通路對機體的影響有待進一步深入研究。