引用本文: 賀濤, 江黎, 程谷萌, 邢怡橋. 5-脂氧合酶對缺氧誘導的視網膜新生血管形成的抑制作用. 中華眼底病雜志, 2014, 30(4): 390-394. doi: 10.3760/cma.j.issn.1005-1015.2014.04.014 復制
視網膜組織缺氧可以引起視網膜新生血管形成,但其形成機制目前還不甚明了。脂氧合酶(LOX)是花生四烯酸代謝過程中重要的限速酶,在各種生理活動中起到重要的作用[1]。根據LOX氧化花生四烯酸時氧插入的不同碳原子位置而分為5-, 8-, 12-, 15-LOX 4種亞型。其中5-LOX具加氧酶和氫過氧化物酶的雙重活性,在人體內分布廣泛,是機體催化花生四烯酸生成生物活性分子,從而影響細胞信號傳導及代謝的關鍵酶。有研究證實,5-LOX參與新生血管的形成并促進炎癥相關腺瘤的形成[2-4]。因此,本研究觀察了5-LOX在視網膜組織中表達,旨在探討5-LOX在缺氧誘導的視網膜新生血管形成過程中可能的作用及機制。現將結果報道如下。
1 材料和方法
C57BL/6J小鼠60只,7日齡,雌雄不限。武漢大學A3實驗動物中心提供。按隨機數字表法分為大劑量干預組、小劑量干預組、干預對照組、氧誘導視網膜病變(OIR)模型組、正常組,各組均為12只。
7日齡C57BL/6J小鼠與哺乳母鼠一同置于氧濃度為(75±2)%的氧箱內。飼養5 d后即小鼠12日齡時取出小鼠轉移至正常環境中飼養5 d,建立OIR模型[5]。12日齡時,大劑量干預組、小劑量干預組小鼠腹腔分別注射100、50 mg/kg 5-LOX抑制劑去甲二氫愈創木酸(NDGA),1次/d,連續5 d;干預對照組小鼠腹腔注射等體積1%二甲基亞砜,1次/d,連續5 d;OIR模型組小鼠出氧箱后不作任何處理;正常組小鼠始終生活在正常氧環境中。
17日齡時處死所有小鼠,摘除眼球。各組分別取12只眼球放入4%多聚甲醛溶液中,4℃,固定24 h。常規酒精梯度脫水,二甲苯透明,石蠟包埋,與視神經矢狀軸平行的連續切片,相鄰2張切片間隔30 μm,每只眼球取切片10張行蘇木精-伊紅(HE)染色,CD34標記新生血管內皮細胞。光學顯微鏡下觀察,雙盲法計數,在400倍鏡下,每張切片取2個視野計數,計算突破內界膜的血管內皮細胞核數。
逆轉錄聚合酶鏈反應(RT-PCR)檢測小鼠視網膜組織5-LOX、血管內皮生長因子(VEGF)-a、VEGF受體-2(VEGFR-2) mRNA的表達。各組分別取12只眼球于手術顯微鏡下去除角膜、晶狀體及玻璃體,完整取出視網膜組織并立即凍存于液氮中備用。采用Trizol一步法提取視網膜組織總RNA。按逆轉錄試劑盒(Toyobo)的操作說明逆轉錄合成cDNA,用以cDNA為模板進行聚合酶鏈反應(PCR)擴增,總反應體系為25.0 μl。引物序列,5-LOX:上游5′-GCTT ATCTGCGAGTATGGC-3′,下游5′-GGATGTCT TCCGTGCTGT-3′,擴增片段大小為158堿基對(bp);VEGF-a:上游5′-TGTCACCACCACGCC ATCA-3′,下游5′-GCCTTTCATCCCATTGTC-3′,擴增片段大小為455 bp;VEGFR-2:上游5′-TCC AAGAAGTGGCAACGG-3′,下游5′-GGAACTCGC CTGTAACCC-3′,擴增片段大小為330 bp;β-肌動蛋白(β-actin):上游5′-CTGTCCCTGTATGCCTCTG-3′,下游5′-ATGTCACGCACGATTTCC-3′,擴增片段大小為218 bp。PCR反應條件:94℃預變性5 min,94℃變性50 s,退火50 s(5-LOX:54.4℃;VEGF-a:59.2℃;VEGFR-2:59.0℃;β-actin:55.0℃),72℃延伸50 s,循環30次后,72℃再延伸10 min。反應后取擴增的PCR產物各4.5 μl,1.5%瓊脂糖凝膠電泳鑒定,電泳完畢后凝膠成像系統下觀察。Bandscan 5.0凝膠軟件分析電泳5-LOX、VEGF-a、VEGFR-2、β-actin的吸光度[A, 舊稱光密度(OD)]值,以β-actin為內參,以每組標本A值的比值評定5-LOX、VEGF-a及VEGFR-2 mRNA的相對表達水平。
蛋白質免疫印跡法(Western blot)檢測5-LOX、VEGF-a、VEGFR-2、細胞外磷酸化信號調節蛋白激酶(P-ERK)1/2的蛋白表達。各組分別取12只眼球于手術顯微鏡下完整取出視網膜組織并立即凍存于液氮中備用。提取視網膜組織蛋白,測定其濃度,10%聚丙烯酰胺凝膠電泳分離蛋白,經80 V電轉2 h,蛋白質印跡轉移到硝酸纖維素膜,然后用洗膜緩沖液(TBST)配置的5%脫脂牛奶封閉2 h,Tris鹽酸緩沖鹽溶液洗滌3次,加一抗(Santa Cruz),濃度分別為5-LOX:1 :500;VEGF-a:1 :500;VEGFR-2:1 :500;P-ERK1/2:1 :500;β-actin:1 :1000。保鮮膜封好4℃過夜后TBST洗滌3次,用TBST按1 :2000的比例稀釋IgG二抗,膜在稀釋液中37℃孵育1 h,最后化學發光顯影,觀察膠片上的條帶并照相保存。采用Bandscan 5.0凝膠分析軟件對特異性條帶進行半定量分析,以5-LOX、VEGF-a、VEGFR-2及P-ERK1/2分別與β-actin的A值的比值評定其蛋白表達水平。
采用SPSS13.0統計軟件包進行統計學分析處理。實驗數據以均數±標準差(
2 結果
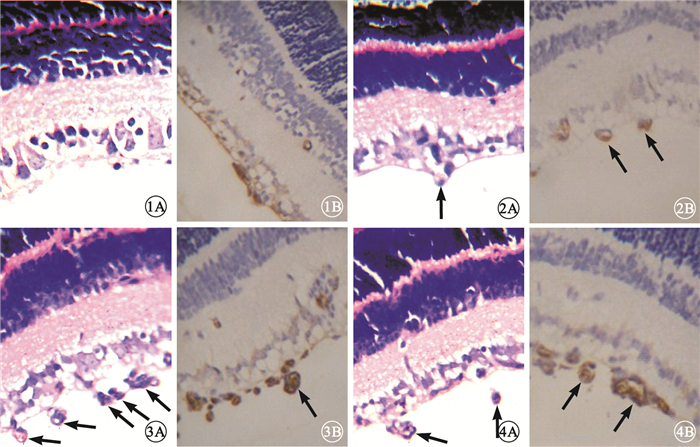
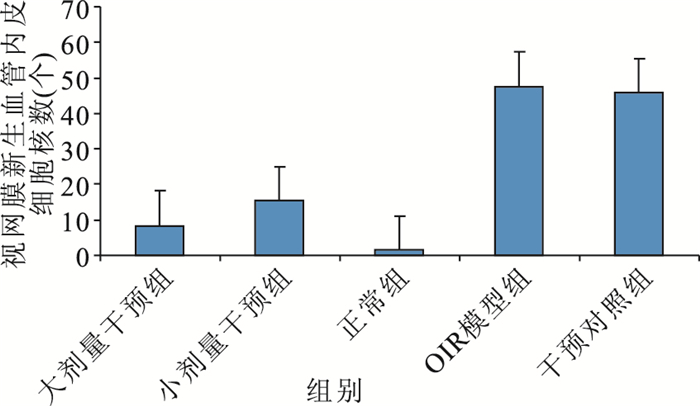
大劑量干預組小鼠視網膜上可見少量新生血管,部分僅為與內界膜相連的單個細胞,視網膜各層排列較為整齊;小劑量干預組小鼠視網膜新生血管稀疏,管腔相對細小,部分尚未形成管腔;正常組小鼠視網膜未見與內界膜相連的到達玻璃體腔的新生血管;OIR模型組、干預對照組小鼠視網膜可見大量突破內界膜伸向玻璃體腔的新生血管,管腔粗大,單獨或成簇出現(圖 1~4)。各組突破內界膜的視網膜新生血管內皮細胞核數比較,差異有統計學意義(F=73.390, P<0.05)(圖 5)。正常組突破內界膜的視網膜新生血管內皮細胞核數與大劑量干預組比較、OIR模型組與干預對照組比較,差異均無統計學意義(F=2.510,2.392,P>0.05);其余各組間比較,差異有統計學意義(P<0.05)。
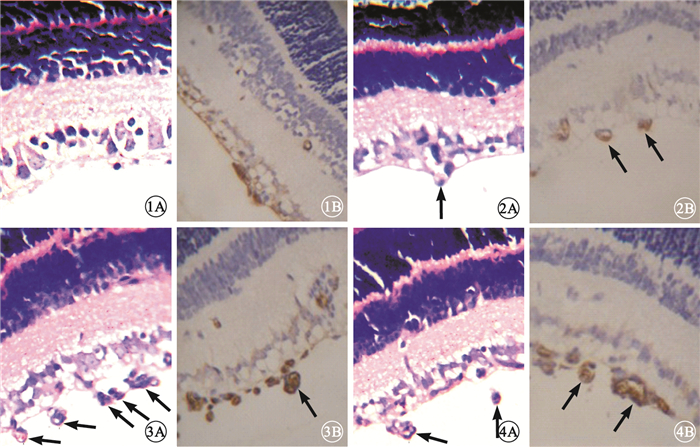 圖1
大劑量組小鼠視網膜組織病理像。1A.未見明顯新生血管,部分僅為與內界膜相連的單個細胞,視網膜各層排列較為整齊??HE ×400;1B.棕黃色特異性著染部分為CD34標記的新生血管壁??HE ×400??圖 2??小劑量組小鼠視網膜組織病理像。2A.新生血管稀疏(黑箭),管腔相對細小,部分尚未形成管腔??HE ×400;2B.CD34標記的新生血管(黑箭)??HE ×400??圖 3??OIR模型組小鼠視網膜組織病理像。3A.可見大量突破內界膜伸向玻璃體腔的新生血管(黑箭)??HE ×400;3B.CD34標記的新生血管(黑箭)??HE ×400??圖 4??干預對照組小鼠視網膜組織病理像。4A.可見大量突破內界膜伸向玻璃體腔的新生血管(黑箭)??HE ×400;4B.CD34標記的新生血管(黑箭)??HE ×400
圖1
大劑量組小鼠視網膜組織病理像。1A.未見明顯新生血管,部分僅為與內界膜相連的單個細胞,視網膜各層排列較為整齊??HE ×400;1B.棕黃色特異性著染部分為CD34標記的新生血管壁??HE ×400??圖 2??小劑量組小鼠視網膜組織病理像。2A.新生血管稀疏(黑箭),管腔相對細小,部分尚未形成管腔??HE ×400;2B.CD34標記的新生血管(黑箭)??HE ×400??圖 3??OIR模型組小鼠視網膜組織病理像。3A.可見大量突破內界膜伸向玻璃體腔的新生血管(黑箭)??HE ×400;3B.CD34標記的新生血管(黑箭)??HE ×400??圖 4??干預對照組小鼠視網膜組織病理像。4A.可見大量突破內界膜伸向玻璃體腔的新生血管(黑箭)??HE ×400;4B.CD34標記的新生血管(黑箭)??HE ×400
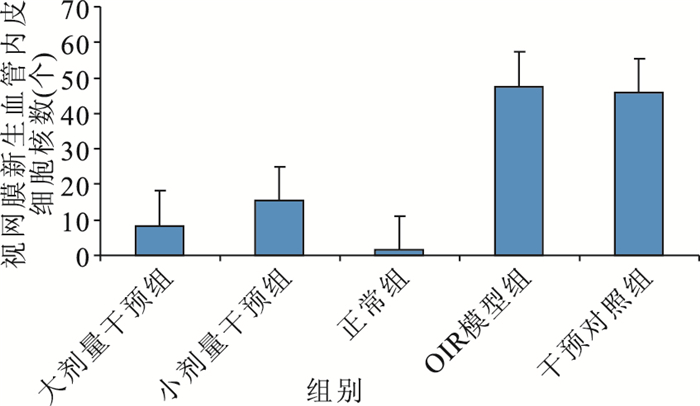 圖5
各組突破內界膜的視網膜新生血管內皮細胞核數比較
圖5
各組突破內界膜的視網膜新生血管內皮細胞核數比較
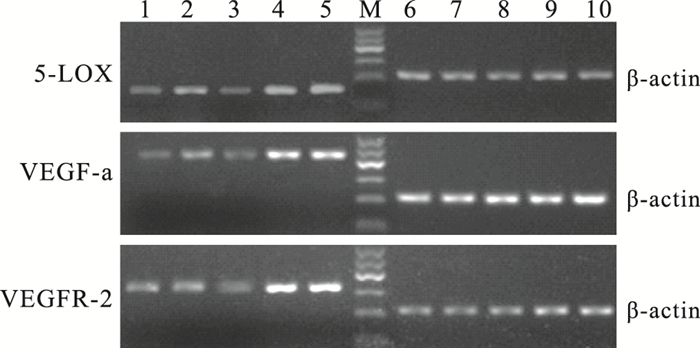
RT-PCR檢測結果顯示,5-LOX mRNA相對表達量OIR模型組小鼠視網膜最高,是正常組、大劑量干預組、小劑量干預組、干預對照組小鼠視網膜的6.29、3.36、1.68、1.03倍。OIR模型組小鼠視網膜5-LOX mRNA相對表達量與正常組、大劑量干預組、小劑量干預組、干預對照組比較,差異有統計學意義(F=92.668,P<0.05)。各組間5-LOX mRNA相對表達量比較,正常組與大劑量干預組、OIR模型組與干預對照組比較,差異無統計學意義(F=1.950、2.331, P>0.05);其余各組比較,差異有統計學意義(P<0.05)(圖 7, 表 1)。
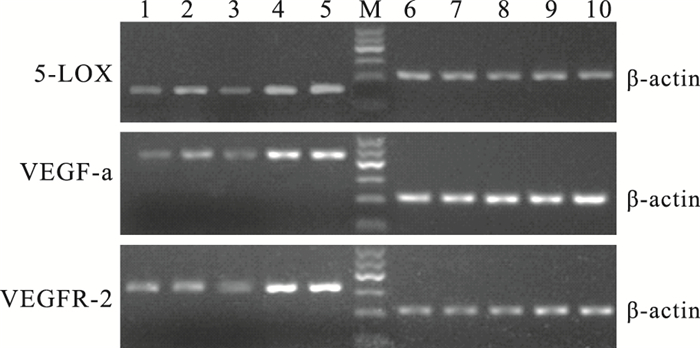 圖7
各組小鼠視網膜組織中5-LOX、VEGF-a及VEGFR-2的mRNA表達電泳圖。1~5:分別為大劑量干預組、小劑量干預組、正常組、OIR模型組、干預對照組;6~10:分別為1~5對應的內參基因β-actin;M:Marker,100~600 bp
圖7
各組小鼠視網膜組織中5-LOX、VEGF-a及VEGFR-2的mRNA表達電泳圖。1~5:分別為大劑量干預組、小劑量干預組、正常組、OIR模型組、干預對照組;6~10:分別為1~5對應的內參基因β-actin;M:Marker,100~600 bp
VEGF-a mRNA相對表達量OIR模型組小鼠視網膜最高,是正常組、大劑量干預組、小劑量干預組、干預對照組的7.00、6.22、2.24、1.03倍。OIR模型組小鼠視網膜VEGF-a mRNA相對表達量與正常組、大劑量干預組、小劑量干預組、干預對照組比較,差異有統計學意義(F=191.390,P<0.05)。各組間VEGF-a mRNA相對表達量比較,正常組與大劑量治療組、OIR模型組與干預對照組比較,差異無統計學意義(F=1.235、2.214, P>0.05);其余各組間比較,差異均有統計學意義(P<0.05)(圖 7,表 1)。
VEGFR-2 mRNA相對表達量OIR模型組小鼠視網膜最高,是正常組、大劑量干預組、小劑量干預組、干預對照組的3.44、1.98、1.89、1.03倍(表 1,圖 7)。OIR模型組小鼠視網膜VEGFR-2mRNA相對表達量與正常組、大劑量干預組、小劑量干預組、干預對照組比較,差異有統計學意義(F=99.604,P<0.05)。各組間VEGFR-2 mRNA相對表達量比較,OIR模型組與干預對照組比較、大劑量干預組與小劑量干預組比較,差異無統計學意義(F=2.512、2.110, P>0.05);其余各組間比較,差異均有統計學意義(P<0.05)(圖 7,表 1)。
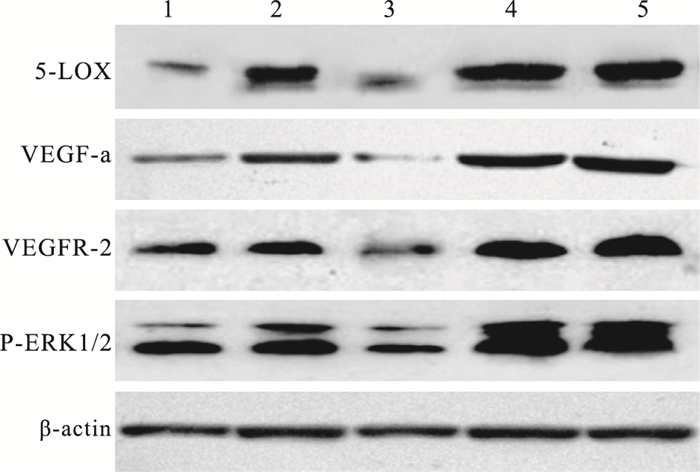
Western blot檢測結果顯示,各組小鼠視網膜中5-LOX、VEGF-a、VEGFR-2、P-ERK1/2蛋白表達具有相同的趨勢,且各組4種蛋白表達量具有顯著差異(F=86.490、165.128、87.092、139.424,P<0.05)。OIR模型組、干預對照組小鼠視網膜各蛋白表達量最高,但兩組間各蛋白表達量比較,差異無統計學意義(F=2.360, P>0.05);正常組各蛋白表達量最低;大劑量干預組各蛋白表達量均低于小劑量干預組,但仍高于正常組各蛋白表達量(表 2,圖 8)。
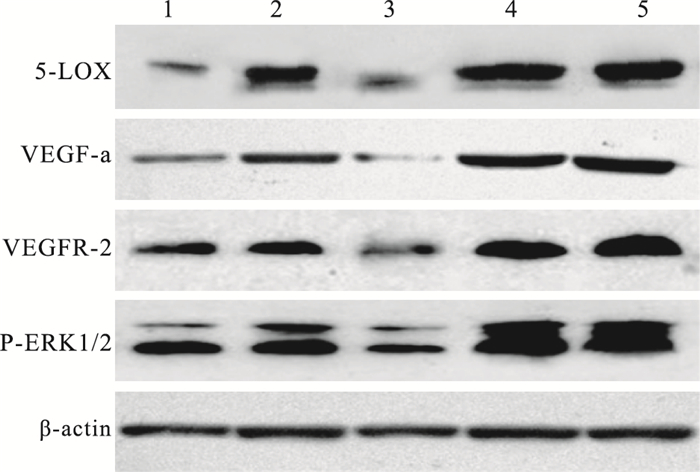 圖8
各組小鼠視網膜組織中5-LOX、VEGFa及VEGFR-2的蛋白質表達電泳圖。1~5:分別為大劑量干預組、小劑量干預組、正常組、OIR模型組、干預對照組
圖8
各組小鼠視網膜組織中5-LOX、VEGFa及VEGFR-2的蛋白質表達電泳圖。1~5:分別為大劑量干預組、小劑量干預組、正常組、OIR模型組、干預對照組
各組間5-LOX蛋白表達量比較,正常組與大劑量治療組、OIR模型組與干預對照組比較,差異無統計學意義(F=1.397、2.005, P>0.05);其余各組間比較,差異均有統計學意義(P<0.05)。VEGF-a蛋白表達量比較,正常組與大劑量治療組、OIR模型組與干預對照組比較,差異無統計學意義(F=1.200、1.959, P>0.05);其余各組間比較,差異均有統計學意義(P<0.05)。VEGFR-2蛋白表達量比較,OIR模型組與干預對照組、大劑量干預組與小劑量干預組比較,差異無統計學意義(F=2.533、2.118, P>0.05);其余各組間比較,差異均有統計學意義(P<0.05)。P-ERK1/2蛋白表達量比較,OIR模型組與干預對照組比較,差異無統計學意義(F=1.990、1.746, P=0.595);其余各組間比較,差異均有統計學意義(P<0.05)。
3 討論
在病理生理代謝過程中占據重要地位的花生四烯酸代謝過程主要有兩種限速酶起到重要的作用即環氧合酶(COX)和LOX[6],其代謝產物參與各種病理生理過程。在LOX的亞家族中的5-LOX是機體催化花生四烯酸生成生物活性分子從而影響細胞信號傳導及代謝的關鍵酶,其催化生成的產物可以通過刺激促血管生成因子如堿性成纖維細胞生長因子(bFGF)以及VEGF等的表達而促進新生血管的生成[7], 為了觀察5-LOX在視網膜新生血管形成過程中的作用機制,本研究建立OIR小鼠模型,應用LOX的選擇性抑制劑NDGA對模型小鼠的5-LOX表達進行抑制,利用RT-PCR及Western blot對5-LOX的mRNA及蛋白質表達進行檢測。結果表明,NDGA可以明顯抑制缺氧誘導的視網膜組織中的5-LOX高表達,與OIR模型組比較,大劑量干預組、小劑量干預組的5-LOX mRNA、蛋白質表達水平均顯著下降,視網膜新生血管形成受到明顯抑制,并呈現明顯的劑量依賴性。說明缺氧誘導的視網膜組織中的5-LOX上調表達與視網膜新生血管的形成有關,用NDGA抑制5-LOX的表達后可以顯著抑制缺氧誘導的視網膜新生血管的形成,從而在惡性腫瘤的發生發展以及轉移中起到重要作用。
在本研究中,OIR模型組較正常組視網膜組織中的VEGF-a mRNA及蛋白質表達上調,說明相對缺氧誘導的VEGF高表達導致了視網膜新生血管的形成。VEGF通過與其特異性受體結合而發揮作用,目前發現的VEGFR主要有VEGFR-1、VEGFR-2及VEGFR-3, 在視網膜組織中主要表達VEGFR-1和VEGFR-2兩種受體,而VEGF的促血管內皮細胞增生和血管形成等生物學功能主要通過VEGFR-2介導[8]。在本研究中,OIR模型組較正常組視網膜組織中的VEGFR-2 mRNA及蛋白質表達上調,說明VEGF和VEGFR-2途徑在病理性血管新生過程中起到了重要的作用。在給予NDGA后,視網膜組織中低氧誘導的VEGF-a和VEGFR-2過量表達被抑制,大劑量干預、小劑量干預組小鼠視網膜中的VEGF-a及VEGFR-2的mRNA及蛋白質表達均較OIR模型組顯著下降,且VEGF-a的表達存在明顯的劑量依賴關系。表明5-LOX可以調節VEGF-a及VEGFR-2的表達,5-LOX受抑制后可以通過減少VEGF-a及VEGFR-2的表達而起到抗血管生成的作用。
ERK1/2是介導血管新生的一類重要的信號蛋白,通過細胞漿或細胞核中其下游的信號分子發揮調節多種內皮細胞遷移、分化和增生的作用[9]。在本研究中,正常組小鼠視網膜中存在P-ERKl/2蛋白質的表達,說明P-ERKl/2參與了視網膜正常生理功能的維持。OIR模型組小鼠視網膜中P-ERKl/2的蛋白質表達相對量較正常組上升3.08倍,表明視網膜組織相對缺氧誘導了P-ERKl/2蛋白質表達的明顯增加。因為磷酸化狀態的ERKl/2為其功能狀態,說明ERKl/2調節通路參與了缺氧誘導的視網膜新生血管的形成過程。干預組小鼠視網膜P-ERKl/2蛋白質表達較模型組明顯下降,且降低程度與抑制劑濃度差異呈現一致性,說明5-LOX抑制導致的視網膜新生血管形成減少與ERKl/2信號的減弱有關系。但是本研究只對缺氧誘導的視網膜新生血管生成過程中P-ERKl/2的表達變化進行了初步觀察,未能闡明P-ERKl/2的調控水平及其對下游底物的作用。因此,尚需在現有研究基礎上對P-ERKl/2在缺氧導致的視網膜新生血管形成過程中的作用進行深入研究以明了其作用機制。
視網膜組織缺氧可以引起視網膜新生血管形成,但其形成機制目前還不甚明了。脂氧合酶(LOX)是花生四烯酸代謝過程中重要的限速酶,在各種生理活動中起到重要的作用[1]。根據LOX氧化花生四烯酸時氧插入的不同碳原子位置而分為5-, 8-, 12-, 15-LOX 4種亞型。其中5-LOX具加氧酶和氫過氧化物酶的雙重活性,在人體內分布廣泛,是機體催化花生四烯酸生成生物活性分子,從而影響細胞信號傳導及代謝的關鍵酶。有研究證實,5-LOX參與新生血管的形成并促進炎癥相關腺瘤的形成[2-4]。因此,本研究觀察了5-LOX在視網膜組織中表達,旨在探討5-LOX在缺氧誘導的視網膜新生血管形成過程中可能的作用及機制。現將結果報道如下。
1 材料和方法
C57BL/6J小鼠60只,7日齡,雌雄不限。武漢大學A3實驗動物中心提供。按隨機數字表法分為大劑量干預組、小劑量干預組、干預對照組、氧誘導視網膜病變(OIR)模型組、正常組,各組均為12只。
7日齡C57BL/6J小鼠與哺乳母鼠一同置于氧濃度為(75±2)%的氧箱內。飼養5 d后即小鼠12日齡時取出小鼠轉移至正常環境中飼養5 d,建立OIR模型[5]。12日齡時,大劑量干預組、小劑量干預組小鼠腹腔分別注射100、50 mg/kg 5-LOX抑制劑去甲二氫愈創木酸(NDGA),1次/d,連續5 d;干預對照組小鼠腹腔注射等體積1%二甲基亞砜,1次/d,連續5 d;OIR模型組小鼠出氧箱后不作任何處理;正常組小鼠始終生活在正常氧環境中。
17日齡時處死所有小鼠,摘除眼球。各組分別取12只眼球放入4%多聚甲醛溶液中,4℃,固定24 h。常規酒精梯度脫水,二甲苯透明,石蠟包埋,與視神經矢狀軸平行的連續切片,相鄰2張切片間隔30 μm,每只眼球取切片10張行蘇木精-伊紅(HE)染色,CD34標記新生血管內皮細胞。光學顯微鏡下觀察,雙盲法計數,在400倍鏡下,每張切片取2個視野計數,計算突破內界膜的血管內皮細胞核數。
逆轉錄聚合酶鏈反應(RT-PCR)檢測小鼠視網膜組織5-LOX、血管內皮生長因子(VEGF)-a、VEGF受體-2(VEGFR-2) mRNA的表達。各組分別取12只眼球于手術顯微鏡下去除角膜、晶狀體及玻璃體,完整取出視網膜組織并立即凍存于液氮中備用。采用Trizol一步法提取視網膜組織總RNA。按逆轉錄試劑盒(Toyobo)的操作說明逆轉錄合成cDNA,用以cDNA為模板進行聚合酶鏈反應(PCR)擴增,總反應體系為25.0 μl。引物序列,5-LOX:上游5′-GCTT ATCTGCGAGTATGGC-3′,下游5′-GGATGTCT TCCGTGCTGT-3′,擴增片段大小為158堿基對(bp);VEGF-a:上游5′-TGTCACCACCACGCC ATCA-3′,下游5′-GCCTTTCATCCCATTGTC-3′,擴增片段大小為455 bp;VEGFR-2:上游5′-TCC AAGAAGTGGCAACGG-3′,下游5′-GGAACTCGC CTGTAACCC-3′,擴增片段大小為330 bp;β-肌動蛋白(β-actin):上游5′-CTGTCCCTGTATGCCTCTG-3′,下游5′-ATGTCACGCACGATTTCC-3′,擴增片段大小為218 bp。PCR反應條件:94℃預變性5 min,94℃變性50 s,退火50 s(5-LOX:54.4℃;VEGF-a:59.2℃;VEGFR-2:59.0℃;β-actin:55.0℃),72℃延伸50 s,循環30次后,72℃再延伸10 min。反應后取擴增的PCR產物各4.5 μl,1.5%瓊脂糖凝膠電泳鑒定,電泳完畢后凝膠成像系統下觀察。Bandscan 5.0凝膠軟件分析電泳5-LOX、VEGF-a、VEGFR-2、β-actin的吸光度[A, 舊稱光密度(OD)]值,以β-actin為內參,以每組標本A值的比值評定5-LOX、VEGF-a及VEGFR-2 mRNA的相對表達水平。
蛋白質免疫印跡法(Western blot)檢測5-LOX、VEGF-a、VEGFR-2、細胞外磷酸化信號調節蛋白激酶(P-ERK)1/2的蛋白表達。各組分別取12只眼球于手術顯微鏡下完整取出視網膜組織并立即凍存于液氮中備用。提取視網膜組織蛋白,測定其濃度,10%聚丙烯酰胺凝膠電泳分離蛋白,經80 V電轉2 h,蛋白質印跡轉移到硝酸纖維素膜,然后用洗膜緩沖液(TBST)配置的5%脫脂牛奶封閉2 h,Tris鹽酸緩沖鹽溶液洗滌3次,加一抗(Santa Cruz),濃度分別為5-LOX:1 :500;VEGF-a:1 :500;VEGFR-2:1 :500;P-ERK1/2:1 :500;β-actin:1 :1000。保鮮膜封好4℃過夜后TBST洗滌3次,用TBST按1 :2000的比例稀釋IgG二抗,膜在稀釋液中37℃孵育1 h,最后化學發光顯影,觀察膠片上的條帶并照相保存。采用Bandscan 5.0凝膠分析軟件對特異性條帶進行半定量分析,以5-LOX、VEGF-a、VEGFR-2及P-ERK1/2分別與β-actin的A值的比值評定其蛋白表達水平。
采用SPSS13.0統計軟件包進行統計學分析處理。實驗數據以均數±標準差(
2 結果
大劑量干預組小鼠視網膜上可見少量新生血管,部分僅為與內界膜相連的單個細胞,視網膜各層排列較為整齊;小劑量干預組小鼠視網膜新生血管稀疏,管腔相對細小,部分尚未形成管腔;正常組小鼠視網膜未見與內界膜相連的到達玻璃體腔的新生血管;OIR模型組、干預對照組小鼠視網膜可見大量突破內界膜伸向玻璃體腔的新生血管,管腔粗大,單獨或成簇出現(圖 1~4)。各組突破內界膜的視網膜新生血管內皮細胞核數比較,差異有統計學意義(F=73.390, P<0.05)(圖 5)。正常組突破內界膜的視網膜新生血管內皮細胞核數與大劑量干預組比較、OIR模型組與干預對照組比較,差異均無統計學意義(F=2.510,2.392,P>0.05);其余各組間比較,差異有統計學意義(P<0.05)。
圖1
大劑量組小鼠視網膜組織病理像。1A.未見明顯新生血管,部分僅為與內界膜相連的單個細胞,視網膜各層排列較為整齊??HE ×400;1B.棕黃色特異性著染部分為CD34標記的新生血管壁??HE ×400??圖 2??小劑量組小鼠視網膜組織病理像。2A.新生血管稀疏(黑箭),管腔相對細小,部分尚未形成管腔??HE ×400;2B.CD34標記的新生血管(黑箭)??HE ×400??圖 3??OIR模型組小鼠視網膜組織病理像。3A.可見大量突破內界膜伸向玻璃體腔的新生血管(黑箭)??HE ×400;3B.CD34標記的新生血管(黑箭)??HE ×400??圖 4??干預對照組小鼠視網膜組織病理像。4A.可見大量突破內界膜伸向玻璃體腔的新生血管(黑箭)??HE ×400;4B.CD34標記的新生血管(黑箭)??HE ×400
圖5
各組突破內界膜的視網膜新生血管內皮細胞核數比較
RT-PCR檢測結果顯示,5-LOX mRNA相對表達量OIR模型組小鼠視網膜最高,是正常組、大劑量干預組、小劑量干預組、干預對照組小鼠視網膜的6.29、3.36、1.68、1.03倍。OIR模型組小鼠視網膜5-LOX mRNA相對表達量與正常組、大劑量干預組、小劑量干預組、干預對照組比較,差異有統計學意義(F=92.668,P<0.05)。各組間5-LOX mRNA相對表達量比較,正常組與大劑量干預組、OIR模型組與干預對照組比較,差異無統計學意義(F=1.950、2.331, P>0.05);其余各組比較,差異有統計學意義(P<0.05)(圖 7, 表 1)。
圖7
各組小鼠視網膜組織中5-LOX、VEGF-a及VEGFR-2的mRNA表達電泳圖。1~5:分別為大劑量干預組、小劑量干預組、正常組、OIR模型組、干預對照組;6~10:分別為1~5對應的內參基因β-actin;M:Marker,100~600 bp
VEGF-a mRNA相對表達量OIR模型組小鼠視網膜最高,是正常組、大劑量干預組、小劑量干預組、干預對照組的7.00、6.22、2.24、1.03倍。OIR模型組小鼠視網膜VEGF-a mRNA相對表達量與正常組、大劑量干預組、小劑量干預組、干預對照組比較,差異有統計學意義(F=191.390,P<0.05)。各組間VEGF-a mRNA相對表達量比較,正常組與大劑量治療組、OIR模型組與干預對照組比較,差異無統計學意義(F=1.235、2.214, P>0.05);其余各組間比較,差異均有統計學意義(P<0.05)(圖 7,表 1)。
VEGFR-2 mRNA相對表達量OIR模型組小鼠視網膜最高,是正常組、大劑量干預組、小劑量干預組、干預對照組的3.44、1.98、1.89、1.03倍(表 1,圖 7)。OIR模型組小鼠視網膜VEGFR-2mRNA相對表達量與正常組、大劑量干預組、小劑量干預組、干預對照組比較,差異有統計學意義(F=99.604,P<0.05)。各組間VEGFR-2 mRNA相對表達量比較,OIR模型組與干預對照組比較、大劑量干預組與小劑量干預組比較,差異無統計學意義(F=2.512、2.110, P>0.05);其余各組間比較,差異均有統計學意義(P<0.05)(圖 7,表 1)。
Western blot檢測結果顯示,各組小鼠視網膜中5-LOX、VEGF-a、VEGFR-2、P-ERK1/2蛋白表達具有相同的趨勢,且各組4種蛋白表達量具有顯著差異(F=86.490、165.128、87.092、139.424,P<0.05)。OIR模型組、干預對照組小鼠視網膜各蛋白表達量最高,但兩組間各蛋白表達量比較,差異無統計學意義(F=2.360, P>0.05);正常組各蛋白表達量最低;大劑量干預組各蛋白表達量均低于小劑量干預組,但仍高于正常組各蛋白表達量(表 2,圖 8)。
圖8
各組小鼠視網膜組織中5-LOX、VEGFa及VEGFR-2的蛋白質表達電泳圖。1~5:分別為大劑量干預組、小劑量干預組、正常組、OIR模型組、干預對照組
各組間5-LOX蛋白表達量比較,正常組與大劑量治療組、OIR模型組與干預對照組比較,差異無統計學意義(F=1.397、2.005, P>0.05);其余各組間比較,差異均有統計學意義(P<0.05)。VEGF-a蛋白表達量比較,正常組與大劑量治療組、OIR模型組與干預對照組比較,差異無統計學意義(F=1.200、1.959, P>0.05);其余各組間比較,差異均有統計學意義(P<0.05)。VEGFR-2蛋白表達量比較,OIR模型組與干預對照組、大劑量干預組與小劑量干預組比較,差異無統計學意義(F=2.533、2.118, P>0.05);其余各組間比較,差異均有統計學意義(P<0.05)。P-ERK1/2蛋白表達量比較,OIR模型組與干預對照組比較,差異無統計學意義(F=1.990、1.746, P=0.595);其余各組間比較,差異均有統計學意義(P<0.05)。
3 討論
在病理生理代謝過程中占據重要地位的花生四烯酸代謝過程主要有兩種限速酶起到重要的作用即環氧合酶(COX)和LOX[6],其代謝產物參與各種病理生理過程。在LOX的亞家族中的5-LOX是機體催化花生四烯酸生成生物活性分子從而影響細胞信號傳導及代謝的關鍵酶,其催化生成的產物可以通過刺激促血管生成因子如堿性成纖維細胞生長因子(bFGF)以及VEGF等的表達而促進新生血管的生成[7], 為了觀察5-LOX在視網膜新生血管形成過程中的作用機制,本研究建立OIR小鼠模型,應用LOX的選擇性抑制劑NDGA對模型小鼠的5-LOX表達進行抑制,利用RT-PCR及Western blot對5-LOX的mRNA及蛋白質表達進行檢測。結果表明,NDGA可以明顯抑制缺氧誘導的視網膜組織中的5-LOX高表達,與OIR模型組比較,大劑量干預組、小劑量干預組的5-LOX mRNA、蛋白質表達水平均顯著下降,視網膜新生血管形成受到明顯抑制,并呈現明顯的劑量依賴性。說明缺氧誘導的視網膜組織中的5-LOX上調表達與視網膜新生血管的形成有關,用NDGA抑制5-LOX的表達后可以顯著抑制缺氧誘導的視網膜新生血管的形成,從而在惡性腫瘤的發生發展以及轉移中起到重要作用。
在本研究中,OIR模型組較正常組視網膜組織中的VEGF-a mRNA及蛋白質表達上調,說明相對缺氧誘導的VEGF高表達導致了視網膜新生血管的形成。VEGF通過與其特異性受體結合而發揮作用,目前發現的VEGFR主要有VEGFR-1、VEGFR-2及VEGFR-3, 在視網膜組織中主要表達VEGFR-1和VEGFR-2兩種受體,而VEGF的促血管內皮細胞增生和血管形成等生物學功能主要通過VEGFR-2介導[8]。在本研究中,OIR模型組較正常組視網膜組織中的VEGFR-2 mRNA及蛋白質表達上調,說明VEGF和VEGFR-2途徑在病理性血管新生過程中起到了重要的作用。在給予NDGA后,視網膜組織中低氧誘導的VEGF-a和VEGFR-2過量表達被抑制,大劑量干預、小劑量干預組小鼠視網膜中的VEGF-a及VEGFR-2的mRNA及蛋白質表達均較OIR模型組顯著下降,且VEGF-a的表達存在明顯的劑量依賴關系。表明5-LOX可以調節VEGF-a及VEGFR-2的表達,5-LOX受抑制后可以通過減少VEGF-a及VEGFR-2的表達而起到抗血管生成的作用。
ERK1/2是介導血管新生的一類重要的信號蛋白,通過細胞漿或細胞核中其下游的信號分子發揮調節多種內皮細胞遷移、分化和增生的作用[9]。在本研究中,正常組小鼠視網膜中存在P-ERKl/2蛋白質的表達,說明P-ERKl/2參與了視網膜正常生理功能的維持。OIR模型組小鼠視網膜中P-ERKl/2的蛋白質表達相對量較正常組上升3.08倍,表明視網膜組織相對缺氧誘導了P-ERKl/2蛋白質表達的明顯增加。因為磷酸化狀態的ERKl/2為其功能狀態,說明ERKl/2調節通路參與了缺氧誘導的視網膜新生血管的形成過程。干預組小鼠視網膜P-ERKl/2蛋白質表達較模型組明顯下降,且降低程度與抑制劑濃度差異呈現一致性,說明5-LOX抑制導致的視網膜新生血管形成減少與ERKl/2信號的減弱有關系。但是本研究只對缺氧誘導的視網膜新生血管生成過程中P-ERKl/2的表達變化進行了初步觀察,未能闡明P-ERKl/2的調控水平及其對下游底物的作用。因此,尚需在現有研究基礎上對P-ERKl/2在缺氧導致的視網膜新生血管形成過程中的作用進行深入研究以明了其作用機制。