引用本文: 黃棋, 田敏, 周琦, 呂紅彬. 5,6-二氫環戊烯1,2-二硫雜環戊烯-3-硫酮對高糖環境下大鼠視網膜Müller細胞的影響. 中華眼底病雜志, 2017, 33(3): 290-294. doi: 10.3760/cma.j.issn.1005-1015.2017.03.016 復制
研究表明,糖尿病視網膜病變(DR)不僅是糖尿病的微血管病變,同時也是神經變性作用的結果[1]。既往文獻報道,高糖誘導的神經元和神經膠質細胞損傷通常發生在微血管損傷之前,其機制可能與氧化應激有關[2]。氧化應激中的活性氧(ROS)是糖尿病急慢性并發癥發生發展的重要物質,可以通過對線粒體結構的破壞從而誘導細胞凋亡[3]。氧化應激通路中的轉錄因子核因子-E2相關因子2(Nrf2)在神經細胞的氧化應激損傷過程中發揮著抗氧化、抗凋亡等作用[4,5]。二項酶誘導劑5,6-二氫環戊烯1,2-二硫雜環戊烯-3-硫酮(CPDT)可激活Nrf2信號通路,促使其下游目標基因血紅素合酶-1(HO-1)的轉錄激活[5]。HO-1及其產物組成重要的內源性保護系統,具有抗氧化、抗炎性損傷等作用。有研究表明,B細胞淋巴瘤/白血病-2(Bcl-2)、Bcl-2相關X蛋白(Bax蛋白) 與細胞凋亡損傷有關[6]。視網膜Müller細胞作為視網膜內最大的神經膠質細胞,橫跨視網膜全層,為神經元提供結構和新陳代謝的支持[7]。因此,本研究通過體外培養Müller細胞,觀察高糖對Müller細胞的影響,探討高糖環境下CPDT對Müller細胞的保護作用及其可能的機制。
1 材料和方法
Sprague Dawley大鼠Müller細胞(上海諾辰生物技術有限公司),胎牛血清(美國Gibco公司),Dulbecco改良Eagle培養基培養液(北京Hyclone公司),CPDT(美國Sigma公司),二喹啉甲酸(BCA)蛋白測定試劑盒(中國Beyotime公司),增強化學發光(ECL)檢測試劑盒(中國凱基生物),兔抗大鼠抗體(美國Abcam公司),Cy3標記山羊抗兔多克隆抗體(美國Abcam公司),辣根過氧化物酶(HRP)羊抗兔和HRP羊抗鼠(美國KPL公司),兔抗磷酸甘油醛脫氫酶(GAPDH)、兔抗Nrf2、鼠抗HO-1(美國Abcam公司),兔抗Bcl-2、兔抗Bax(美國Abcam公司)。酶標儀(美國RD公司),電泳儀(美國RD公司)。
免疫熒光染色鑒定Müller細胞。將細胞接種至12孔板中的蓋玻片上,5% CO2培養箱37 ℃培養24 h;取出爬片,加入4%多聚甲醛室溫固定10 min;磷酸鹽緩沖液(PBS)浸洗3次,5 min/次。與一抗兔抗大鼠波形蛋白抗體(1:200)37 ℃反應2 h。PBS浸洗3次,5 min/次。與對應的Cy3標記的山羊抗兔多克隆抗體二抗(1:1000)37 ℃反應1 h。PBS浸洗3次,5 min/次,與hoechst熒光染料反應15 min。PBS浸洗3次,5 min/次,將細胞爬片封片于載玻片拍照。空白對照以PBS代替一抗其余步驟完全相同,計算染色呈陽性細胞數與細胞總數比例,以細胞核計算總數。
參照文獻[8]方法根據葡萄糖濃度及CPDT濃度將Müller細胞分為25 mmol/L對照組(A組)、65 mmol/L高糖組(B組)組、高糖+45 μmol/L CPDT組(C組)、高糖+60 μmol/L CPDT組(D組)、高糖+70 μmol/L CPDT組(E組)。按照分組條件處理各組Müller細胞,5% CO2培養箱37 ℃培養72 h。
取對數生長期Müller細胞,0.25%胰蛋白酶消化單細胞,重懸于新鮮生長培養液中。將細胞按2×103接種于96孔培養板1 h,450 nm波長酶標儀檢測各孔Müller細胞吸光度[A,舊稱光密度(OD)]值。水溶性四唑鹽(WST)-8法檢測Müller細胞相對增生率,重復3次取平均值。
各組細胞培養72 h后收集細胞,流式細胞儀分別檢測A、B、D組Müller細胞中ROS含量及細胞凋亡情況。將培養的第3代細胞消化離心獲得沉淀,加入適量濃度為10 μmol/L的二氯二氫熒光素-乙酰乙酸酯(DCFH-DA),體積以能充分蓋住Müller細胞為宜。37 ℃細胞培養箱內避光孵育30 min。無血清培養液洗滌Müller細胞1~2次,上機檢測。空白對照以等量PBS代替DCFH-DA工作液。Müller細胞消化離心,沉淀收集于流式管中,沉淀用300 μl結合緩沖液重懸。加入5 μl膜聯蛋白V聚乙烯(AnnexinV-PE)混勻后,加入5 μl 7-氨基放線菌素D(7-AAD)混勻。室溫下避光反應15 min。上機檢測。空白對照以等量PBS代替AnnexinV-PE。
蛋白免疫印跡法(Western Blot)檢測A、B、D組Müller細胞中Nrf2、HO-1、Bcl-2和Bax蛋白表達。裂解細胞,提取細胞總蛋白,BCA蛋白測定試劑盒測定蛋白濃度。取30 μg蛋白進行8%十二烷基硫酸鈉聚丙烯酰胺凝膠電泳變性后,將蛋白轉至聚偏二氟乙烯膜。5%脫脂牛奶封閉2 h,一抗4℃孵育過夜,洗膜。常溫孵育二抗1 h,洗膜。ECL檢測試劑盒發光顯影。以GAPDH為內參,計算各組Müller細胞中Nrf2、HO-1、Bcl-2、Bax蛋白表達量。
采用SPSS 20.0統計軟件進行統計學分析處理。數據以均數±標準差( )表示。兩樣本均數比較采用t檢驗,并進行方差齊性檢驗。α=0.05。P<0.05為差異有統計學意義。
2 結果
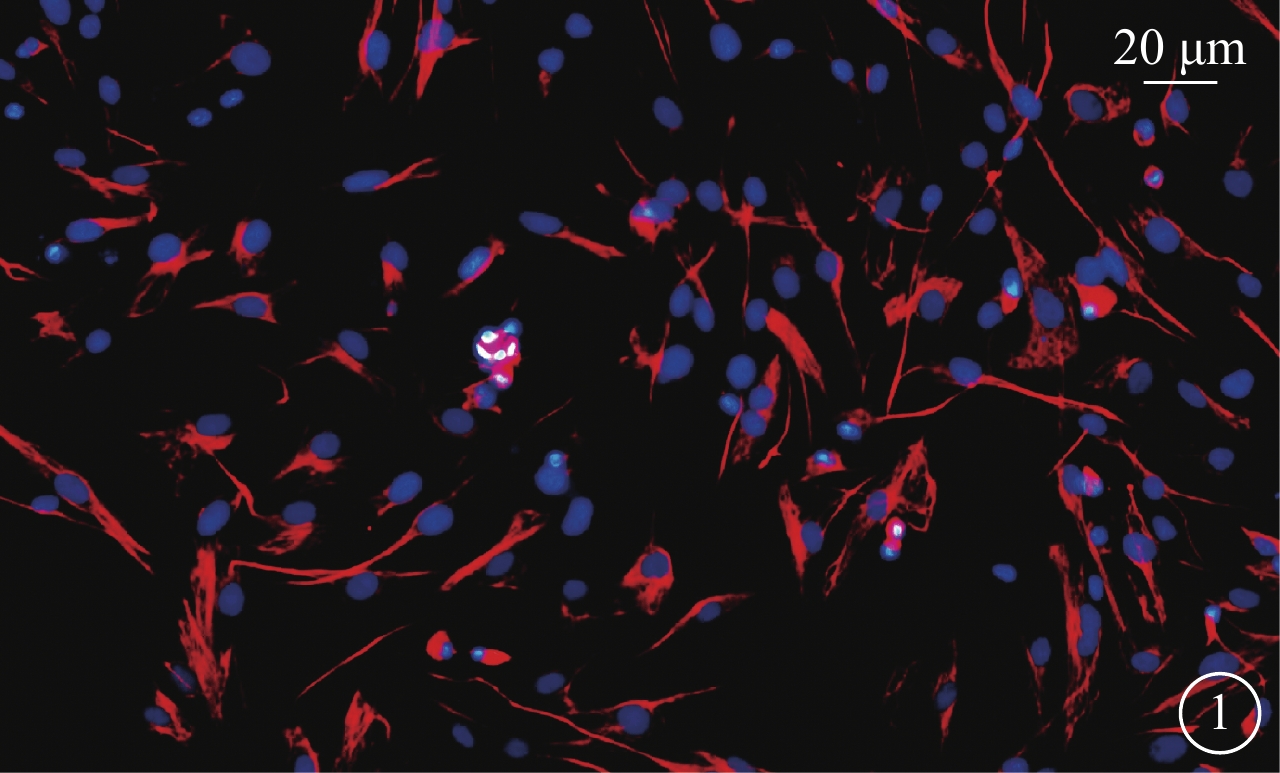
免疫熒光雙標法鑒定結果顯示,Müller細胞抗波形蛋白呈陽性的細胞質可見橘紅色熒光(圖1);Müller細胞陽性率90%以上。倒置熒光顯微鏡觀察,可見Müller細胞胞體肥大、扁平,向四周伸出突起,細胞核呈卵圓形,相鄰細胞交織成網狀,形態基本一致。
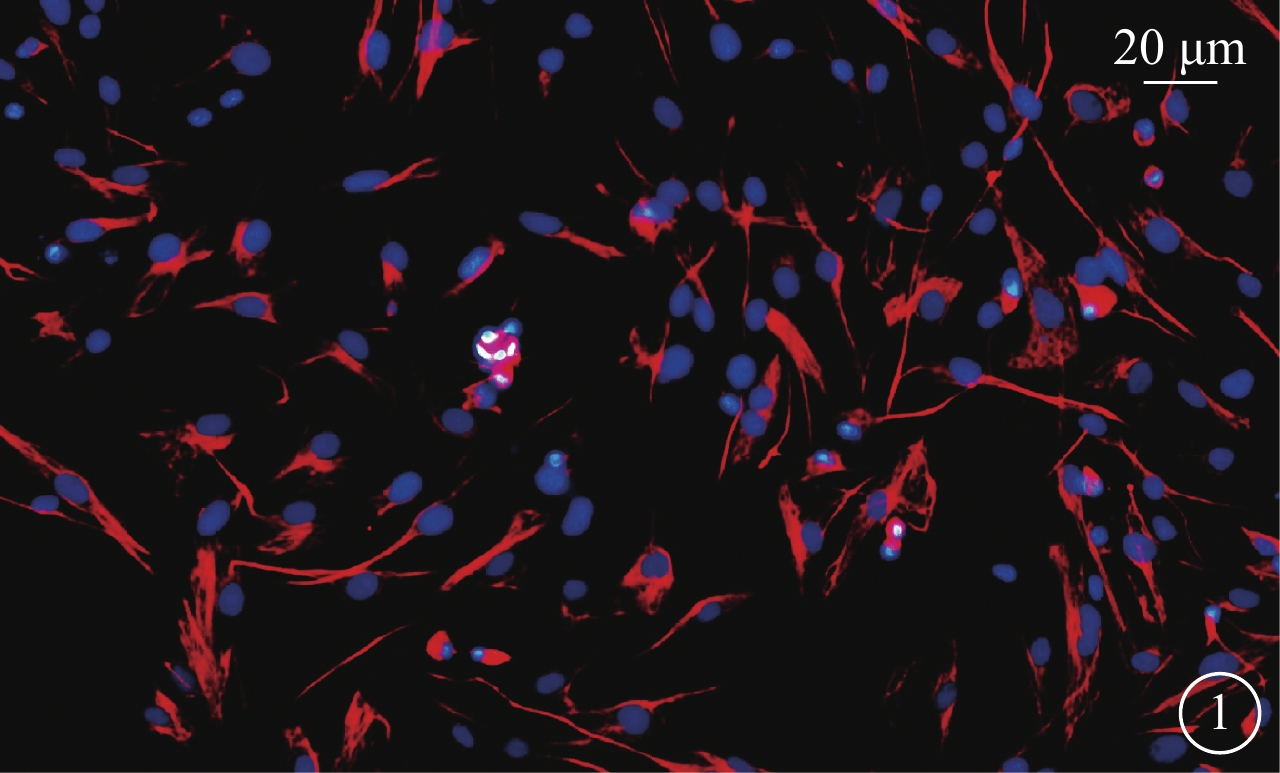 圖1
Müller細胞免疫熒光染色倒置熒光顯微鏡像。Müller細胞細胞質呈橘紅色,細胞核呈藍色 免疫熒光染色 標尺:20 μm
圖1
Müller細胞免疫熒光染色倒置熒光顯微鏡像。Müller細胞細胞質呈橘紅色,細胞核呈藍色 免疫熒光染色 標尺:20 μm
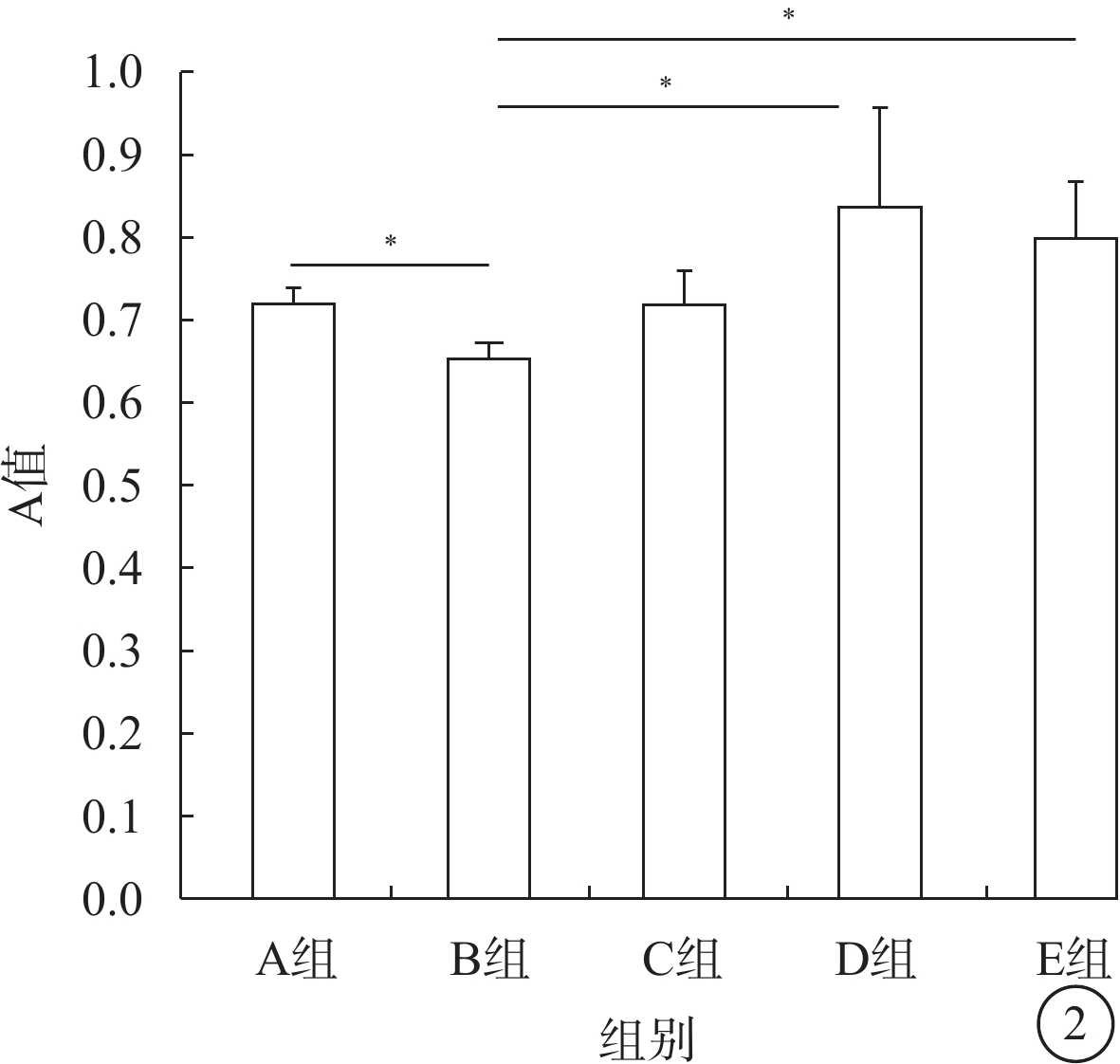
WST-8法檢測結果顯示,B組Müller細胞平均A值較A組低,差異有統計學意義(t=39.59,P<0.05);D組Müller細胞平均A值較B組高,差異有統計學意義(t=?4.17,P<0.05),Müller細胞增生率為155%;E組Müller細胞平均A值較B組高,差異有統計學意義(t=?7.52,P<0.05),Müller細胞增生率為143%;C組Müller細胞平均A值與B組比較,差異無統計學意義(t=0.97,P>0.05)(圖2)。
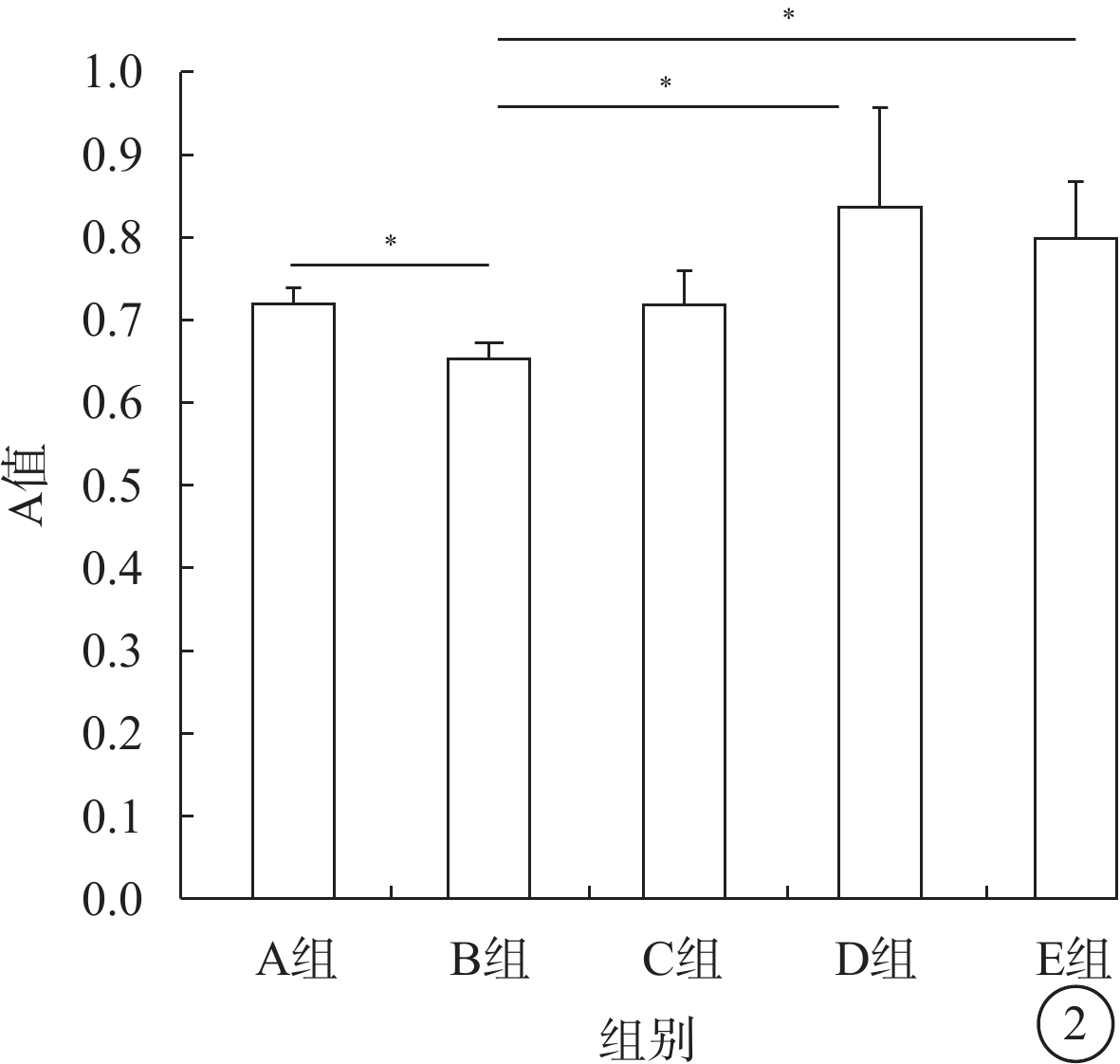 圖2
各組Müller細胞A值比較。*P<0.05
圖2
各組Müller細胞A值比較。*P<0.05
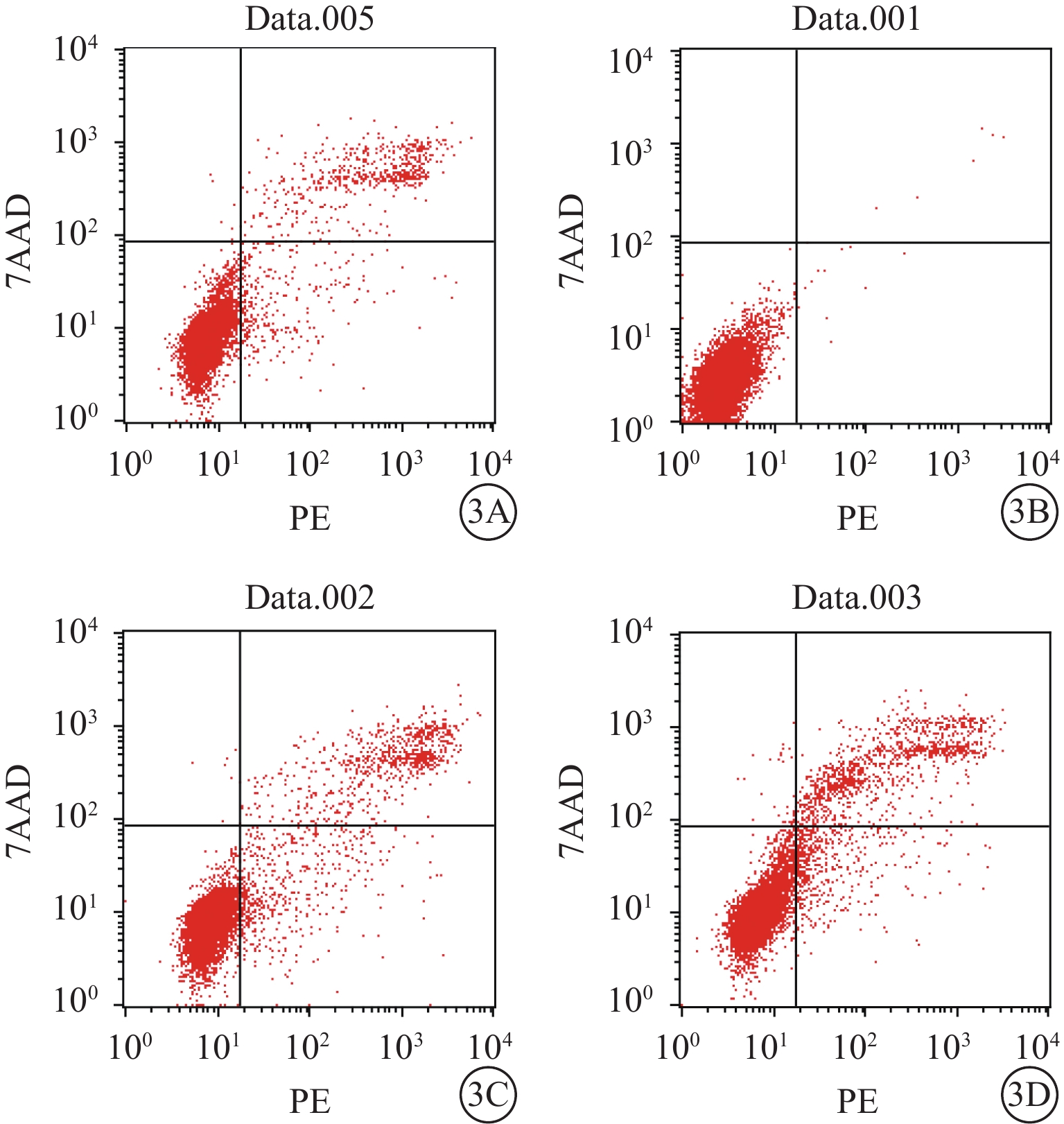
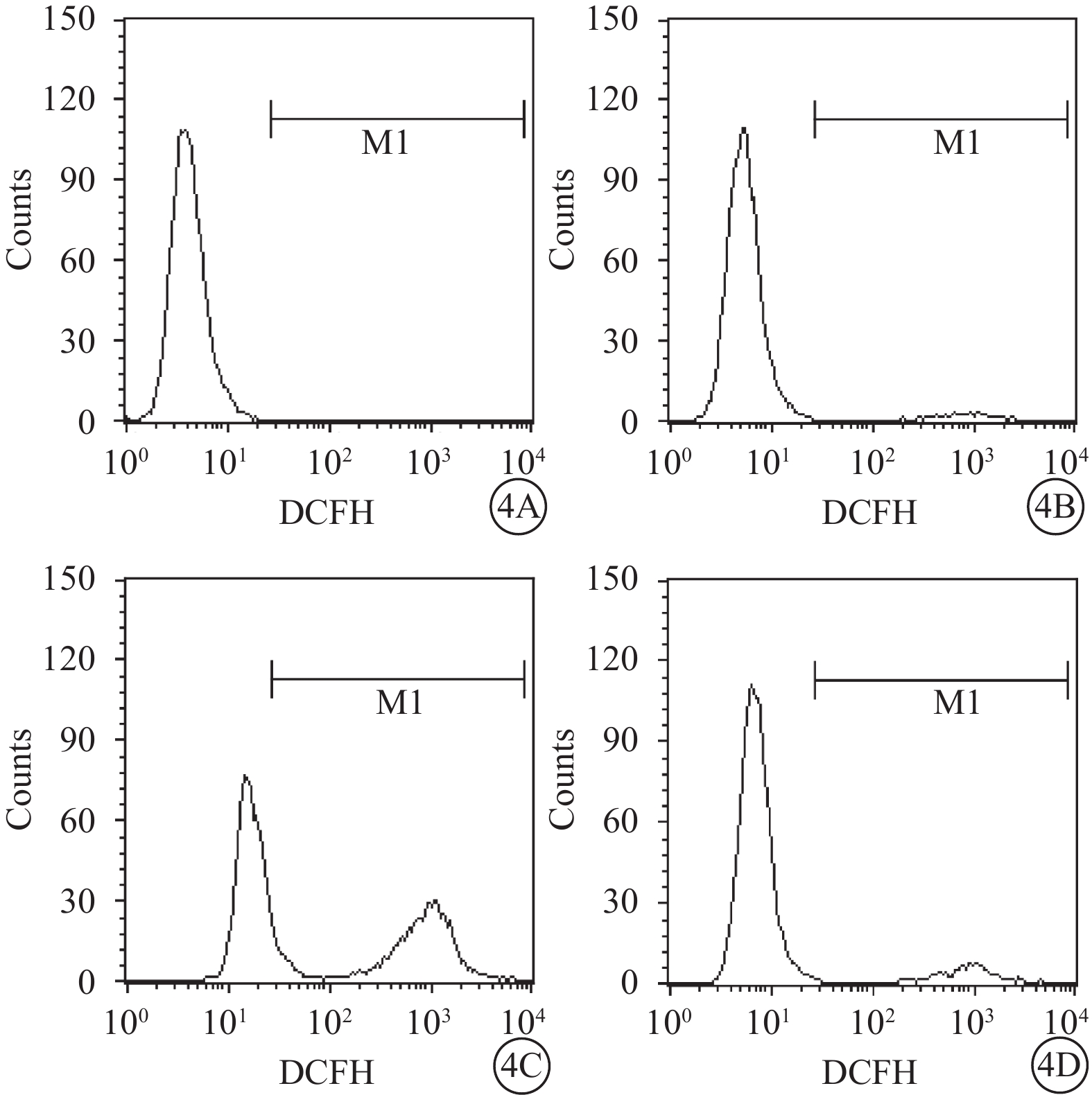
流式細胞儀檢測結果顯示,空白對照Müller細胞凋亡率為0.06%。A、B、D組Müller細胞凋亡率分別為(8.25±0.26)%、(17.60±3.16)%、(13.50±2.25)%;A、B組Müller細胞凋亡率比較,差異有統計學意義(t=?8.91,P<0.05);D組Müller細胞凋亡率較B組降低,差異有統計學意義(t=19.03,P<0.05)(圖3)。空白對照Müller細胞中ROS含量為0.02%。A、B、D組Müller細胞中ROS含量分別3.09%、46.01%、8.01%。與A組比較,B組Müller細胞中ROS含量明顯增多,差異有統計學意義(t=?30.99,P<0.05);與B組比較,D組Müller細胞中ROS含量明顯下降,差異有統計學意義(t=27.68,P<0.05)(圖4)。
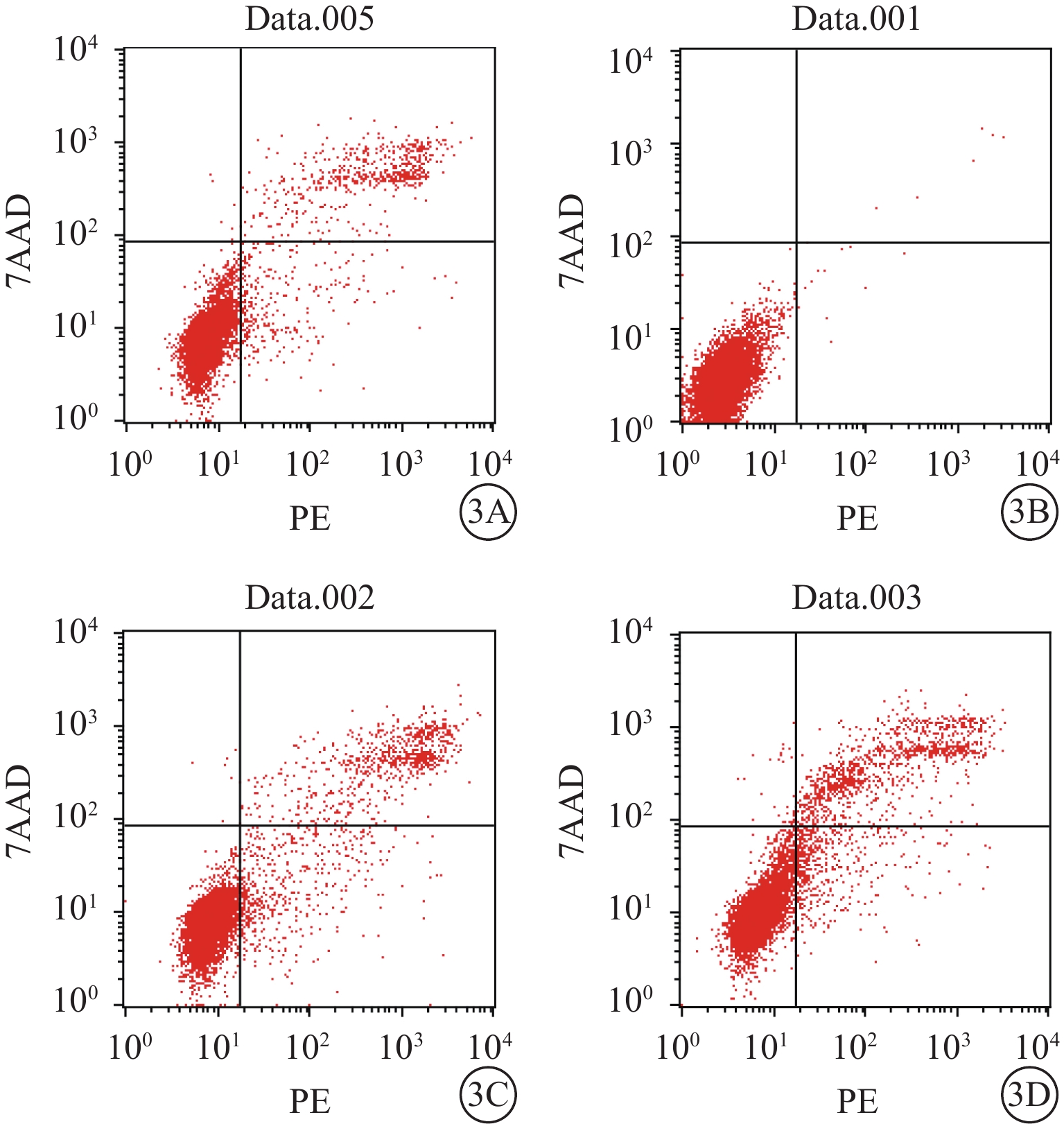 圖3
A、B、D組Müller細胞凋亡率檢測結果。3A~3D. 分別為空白對照,A、B、D組
圖3
A、B、D組Müller細胞凋亡率檢測結果。3A~3D. 分別為空白對照,A、B、D組
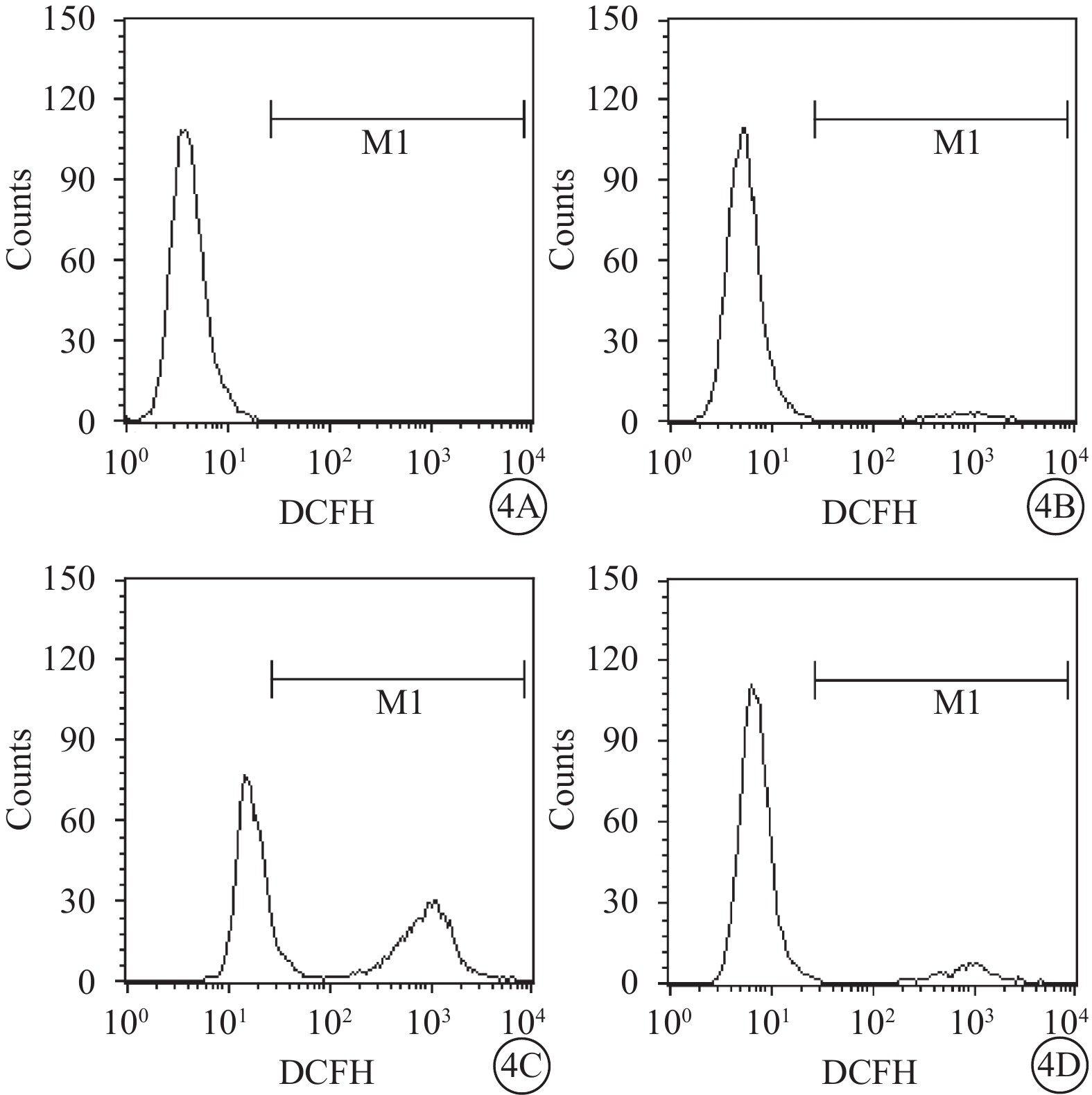 圖4
A、B、D組Müller細胞中ROS含量檢測結果。4A~4D. 分別為空白對照,A、B、D組
圖4
A、B、D組Müller細胞中ROS含量檢測結果。4A~4D. 分別為空白對照,A、B、D組
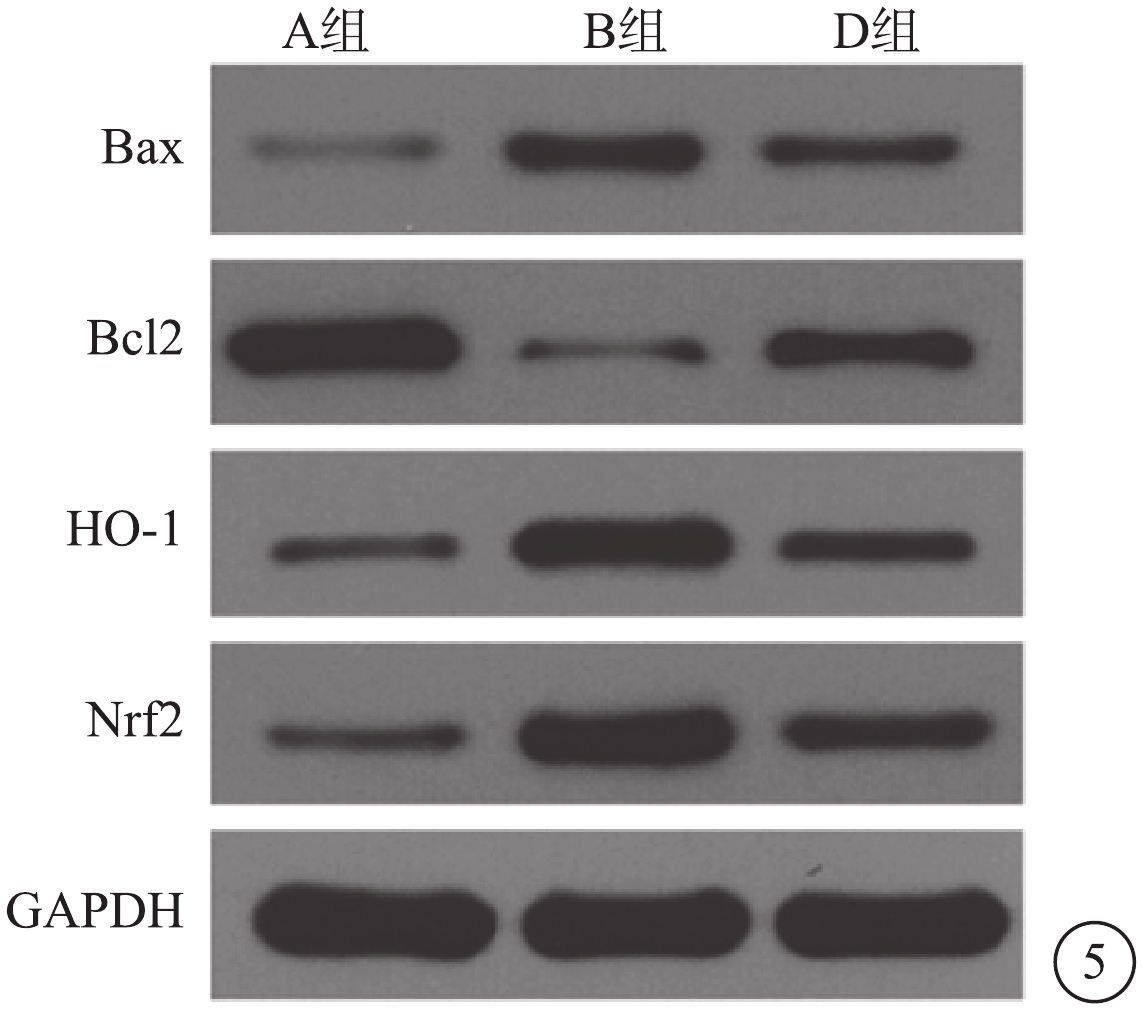
Western blot檢測結果顯示,與A組比較,B組Müller細胞Bax、Nrf2、HO-1蛋白表達顯著上調(t=?11.03、?63.17、?11.44),差異有統計學意義(P<0.05);Bcl-2蛋白表達明顯下調,差異有統計學意義(t=7.86,P<0.05)。與B組比較,D組Müller細胞Nrf2、HO-1、Bax蛋白表達均下調(t=15.11、26.59、6.27),差異有統計學意義(P<0.05); Bcl-2 蛋白表達上調,差異有統計學意義(t=?6.53,P<0.05)(圖5)。與A組比較,B組Müller細胞Bcl-2/Bax比值降低,差異有統計學意義(t=4.84,P<0.05);與B組比較,不同濃度CPDT干預組Müller細胞Bcl-2/Bax比值增高,差異有統計學意義(t=?7.813,P<0.05)。
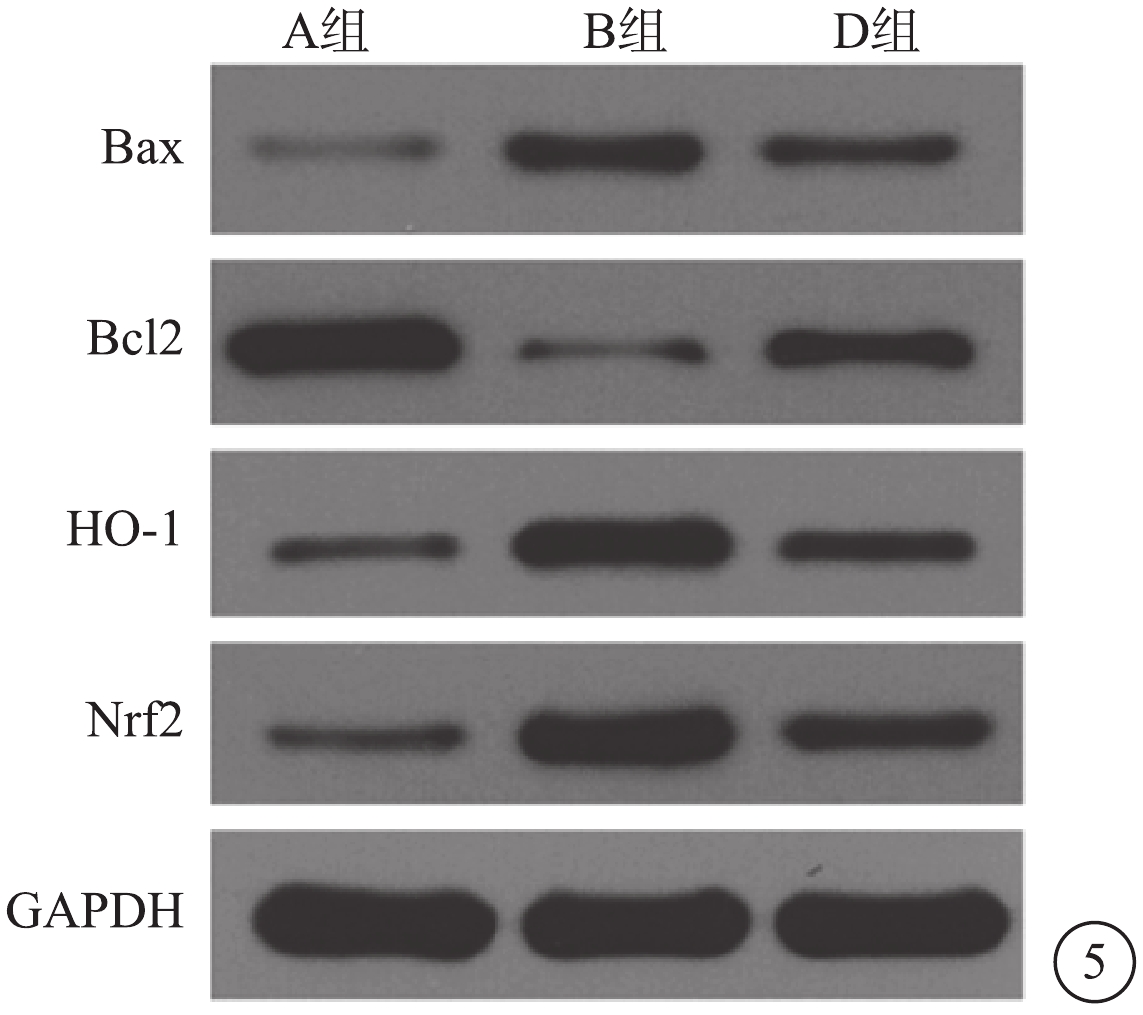 圖5
A、B、D組Müller細胞HO-1、Nrf2、Bcl-2、Bax蛋白表達電泳圖
圖5
A、B、D組Müller細胞HO-1、Nrf2、Bcl-2、Bax蛋白表達電泳圖
3 討論
CPDT作為二項酶誘導劑,已被證實可在神經元中通過誘導二項酶表達對神經元細胞起保護作用。在肌萎縮側索硬化、腦卒中領域作為誘導二項酶及其相關抗氧化酶發揮其抗氧化應激作用。
DR發生時Müller細胞微環境發生改變,其周圍糖濃度增加,此時周圍的葡萄糖及其自身中間產物產生大量如氧自由基和H2O2等ROS,這些氧自由基可以作用于線粒體通路,對線粒體造成損傷。氧化應激損傷的反應主要通過Nrf2/HO-1通路實現,其中,HO-1可促進代謝性炎癥反應和胰島素抵抗等,糖尿病的發生常伴隨HO-1的增高[9-13]。糖尿病發生時也常伴隨多種器官組織的凋亡。細胞凋亡的線粒體途徑通過Bcl-2在線粒體膜外發揮作用維持線粒體膜的完整性,相反,Bax則破壞線粒體膜完整性,當二者失去平衡時,Bcl-2對抗Bax作用減弱,從而誘導凋亡的發生。作為二項酶誘導劑CPDT可以明顯減輕高糖對Müller細胞的氧化應激作用,并能有效抑制Müller細胞凋亡[11-14]。本研究結果發現,與B組相比較,C、D、E組Müller細胞Nrf2、HO-1、Bax蛋白表達均下調,Bcl-2蛋白表達量增加,表明二項酶誘導劑可通過Nrf2/HO-1通路保護Müller細胞,并抑制細胞的凋亡。另有研究表明二項酶誘導劑對2型糖尿病大鼠視網膜損傷也具有一定保護作用[15]。
本研究結果表明,CPDT可能一方面通過激活Nrf2/HO-1氧化應激通路,另一方面增加Bcl-2蛋白表達量,減少線粒體氧自由基量的生成,從而抑制Müller細胞凋亡。隨著基因技術的發展,可利用基因技術對CPDT在高糖條件中的作用及其機制進行深入研究。
讀者?作者?編者 關于本刊財務收支過程中涉及到的財務管理規定及注意事項說明 本刊財務收支均由承辦單位四川大學華西醫院統一管理。華西醫院賬戶眾多收款項目中,期刊收支項目占比較小,安排的工作人員有限,但管理規定及執行程序同樣非常嚴格。為了提高效率,方便工作,現將版面費等支付過程中涉及到的財務管理規定及注意事項說明如下: 稿件送修并確認刊載后本刊將通知作者按要求數額支付版面費。版面費可通過郵局或銀行支付。 郵局支付者,收款人請寫“編輯部”,不能寄給任何個人。匯款單附言欄必須注明開具發票的抬頭和刊用稿件的編號。發票抬頭若開具單位名稱,應以單位組織機構代碼中的規范名稱為準,如四川大學華西醫院。 通過銀行匯款的版面費均匯至四川大學華西醫院銀行賬戶。匯款者若為個人,應在匯款后當天將匯款憑證掃描或照相后發到編輯部(Email:coretina@163.com);若通過單位對公賬戶匯款,請在提交匯款申請后當日將匯款申請信息告之編輯部,1周內將匯款憑證掃描或照相后發到編輯部。以便及時查找確認并開具發票。若未能按要求時間完成上述工作,發票開具時間則不能確定。跨年的轉款項目財務部門原則上不再查賬并開具發票。 發票出具時,匯款者若為個人,發票抬頭只能開具匯款者姓名;發票抬頭若需開具單位名稱,則只能通過發票抬頭單位對公賬戶匯款。凡依據付款方要求出具一次發票后,不再重新開具任何需要修改發票抬頭等內容的發票。收到版面費后3個月內,以快遞方式寄出版面費發票。 版面費收取及發票查詢的聯系方式,聯系人:萬苗,電話:028-85422060。 除版面費外,其他與本刊相關的財務收支及發票出具注意事項參照上述規定和要求執行。 關于作者與編輯部之間通訊聯系的說明 作者投稿均請通過本刊網站完成,本刊不接受電子郵件或郵寄稿件等其他方式投稿。查詢稿件處理進程、了解說明稿件情況等稿件處理相關的所有溝通聯系均請通過編輯部郵箱coretina@163.com進行。投遞給編輯部的各類信函、匯款均應署名“中華眼底病雜志編輯部”收。上述與工作相關的郵件、信函、匯款均不宜發送、投寄給個人,為便于留痕備查。 編輯部稿件處理流程中相關的所有事宜均同時給第一作者和通信作者聯系,重要事宜應由通信作者承擔責任并負責回復。雜志出版后當期雜志、文章單行本以及稿酬、發票等均只郵寄給第一作者;其第一作者投稿后各種聯系方式有變更者,應及時告知編輯部,以免造成相關工作延誤或不可彌補的損失。 本刊編輯部
研究表明,糖尿病視網膜病變(DR)不僅是糖尿病的微血管病變,同時也是神經變性作用的結果[1]。既往文獻報道,高糖誘導的神經元和神經膠質細胞損傷通常發生在微血管損傷之前,其機制可能與氧化應激有關[2]。氧化應激中的活性氧(ROS)是糖尿病急慢性并發癥發生發展的重要物質,可以通過對線粒體結構的破壞從而誘導細胞凋亡[3]。氧化應激通路中的轉錄因子核因子-E2相關因子2(Nrf2)在神經細胞的氧化應激損傷過程中發揮著抗氧化、抗凋亡等作用[4,5]。二項酶誘導劑5,6-二氫環戊烯1,2-二硫雜環戊烯-3-硫酮(CPDT)可激活Nrf2信號通路,促使其下游目標基因血紅素合酶-1(HO-1)的轉錄激活[5]。HO-1及其產物組成重要的內源性保護系統,具有抗氧化、抗炎性損傷等作用。有研究表明,B細胞淋巴瘤/白血病-2(Bcl-2)、Bcl-2相關X蛋白(Bax蛋白) 與細胞凋亡損傷有關[6]。視網膜Müller細胞作為視網膜內最大的神經膠質細胞,橫跨視網膜全層,為神經元提供結構和新陳代謝的支持[7]。因此,本研究通過體外培養Müller細胞,觀察高糖對Müller細胞的影響,探討高糖環境下CPDT對Müller細胞的保護作用及其可能的機制。
1 材料和方法
Sprague Dawley大鼠Müller細胞(上海諾辰生物技術有限公司),胎牛血清(美國Gibco公司),Dulbecco改良Eagle培養基培養液(北京Hyclone公司),CPDT(美國Sigma公司),二喹啉甲酸(BCA)蛋白測定試劑盒(中國Beyotime公司),增強化學發光(ECL)檢測試劑盒(中國凱基生物),兔抗大鼠抗體(美國Abcam公司),Cy3標記山羊抗兔多克隆抗體(美國Abcam公司),辣根過氧化物酶(HRP)羊抗兔和HRP羊抗鼠(美國KPL公司),兔抗磷酸甘油醛脫氫酶(GAPDH)、兔抗Nrf2、鼠抗HO-1(美國Abcam公司),兔抗Bcl-2、兔抗Bax(美國Abcam公司)。酶標儀(美國RD公司),電泳儀(美國RD公司)。
免疫熒光染色鑒定Müller細胞。將細胞接種至12孔板中的蓋玻片上,5% CO2培養箱37 ℃培養24 h;取出爬片,加入4%多聚甲醛室溫固定10 min;磷酸鹽緩沖液(PBS)浸洗3次,5 min/次。與一抗兔抗大鼠波形蛋白抗體(1:200)37 ℃反應2 h。PBS浸洗3次,5 min/次。與對應的Cy3標記的山羊抗兔多克隆抗體二抗(1:1000)37 ℃反應1 h。PBS浸洗3次,5 min/次,與hoechst熒光染料反應15 min。PBS浸洗3次,5 min/次,將細胞爬片封片于載玻片拍照。空白對照以PBS代替一抗其余步驟完全相同,計算染色呈陽性細胞數與細胞總數比例,以細胞核計算總數。
參照文獻[8]方法根據葡萄糖濃度及CPDT濃度將Müller細胞分為25 mmol/L對照組(A組)、65 mmol/L高糖組(B組)組、高糖+45 μmol/L CPDT組(C組)、高糖+60 μmol/L CPDT組(D組)、高糖+70 μmol/L CPDT組(E組)。按照分組條件處理各組Müller細胞,5% CO2培養箱37 ℃培養72 h。
取對數生長期Müller細胞,0.25%胰蛋白酶消化單細胞,重懸于新鮮生長培養液中。將細胞按2×103接種于96孔培養板1 h,450 nm波長酶標儀檢測各孔Müller細胞吸光度[A,舊稱光密度(OD)]值。水溶性四唑鹽(WST)-8法檢測Müller細胞相對增生率,重復3次取平均值。
各組細胞培養72 h后收集細胞,流式細胞儀分別檢測A、B、D組Müller細胞中ROS含量及細胞凋亡情況。將培養的第3代細胞消化離心獲得沉淀,加入適量濃度為10 μmol/L的二氯二氫熒光素-乙酰乙酸酯(DCFH-DA),體積以能充分蓋住Müller細胞為宜。37 ℃細胞培養箱內避光孵育30 min。無血清培養液洗滌Müller細胞1~2次,上機檢測。空白對照以等量PBS代替DCFH-DA工作液。Müller細胞消化離心,沉淀收集于流式管中,沉淀用300 μl結合緩沖液重懸。加入5 μl膜聯蛋白V聚乙烯(AnnexinV-PE)混勻后,加入5 μl 7-氨基放線菌素D(7-AAD)混勻。室溫下避光反應15 min。上機檢測。空白對照以等量PBS代替AnnexinV-PE。
蛋白免疫印跡法(Western Blot)檢測A、B、D組Müller細胞中Nrf2、HO-1、Bcl-2和Bax蛋白表達。裂解細胞,提取細胞總蛋白,BCA蛋白測定試劑盒測定蛋白濃度。取30 μg蛋白進行8%十二烷基硫酸鈉聚丙烯酰胺凝膠電泳變性后,將蛋白轉至聚偏二氟乙烯膜。5%脫脂牛奶封閉2 h,一抗4℃孵育過夜,洗膜。常溫孵育二抗1 h,洗膜。ECL檢測試劑盒發光顯影。以GAPDH為內參,計算各組Müller細胞中Nrf2、HO-1、Bcl-2、Bax蛋白表達量。
采用SPSS 20.0統計軟件進行統計學分析處理。數據以均數±標準差( )表示。兩樣本均數比較采用t檢驗,并進行方差齊性檢驗。α=0.05。P<0.05為差異有統計學意義。
2 結果
免疫熒光雙標法鑒定結果顯示,Müller細胞抗波形蛋白呈陽性的細胞質可見橘紅色熒光(圖1);Müller細胞陽性率90%以上。倒置熒光顯微鏡觀察,可見Müller細胞胞體肥大、扁平,向四周伸出突起,細胞核呈卵圓形,相鄰細胞交織成網狀,形態基本一致。
圖1
Müller細胞免疫熒光染色倒置熒光顯微鏡像。Müller細胞細胞質呈橘紅色,細胞核呈藍色 免疫熒光染色 標尺:20 μm
WST-8法檢測結果顯示,B組Müller細胞平均A值較A組低,差異有統計學意義(t=39.59,P<0.05);D組Müller細胞平均A值較B組高,差異有統計學意義(t=?4.17,P<0.05),Müller細胞增生率為155%;E組Müller細胞平均A值較B組高,差異有統計學意義(t=?7.52,P<0.05),Müller細胞增生率為143%;C組Müller細胞平均A值與B組比較,差異無統計學意義(t=0.97,P>0.05)(圖2)。
圖2
各組Müller細胞A值比較。*P<0.05
流式細胞儀檢測結果顯示,空白對照Müller細胞凋亡率為0.06%。A、B、D組Müller細胞凋亡率分別為(8.25±0.26)%、(17.60±3.16)%、(13.50±2.25)%;A、B組Müller細胞凋亡率比較,差異有統計學意義(t=?8.91,P<0.05);D組Müller細胞凋亡率較B組降低,差異有統計學意義(t=19.03,P<0.05)(圖3)。空白對照Müller細胞中ROS含量為0.02%。A、B、D組Müller細胞中ROS含量分別3.09%、46.01%、8.01%。與A組比較,B組Müller細胞中ROS含量明顯增多,差異有統計學意義(t=?30.99,P<0.05);與B組比較,D組Müller細胞中ROS含量明顯下降,差異有統計學意義(t=27.68,P<0.05)(圖4)。
圖3
A、B、D組Müller細胞凋亡率檢測結果。3A~3D. 分別為空白對照,A、B、D組
圖4
A、B、D組Müller細胞中ROS含量檢測結果。4A~4D. 分別為空白對照,A、B、D組
Western blot檢測結果顯示,與A組比較,B組Müller細胞Bax、Nrf2、HO-1蛋白表達顯著上調(t=?11.03、?63.17、?11.44),差異有統計學意義(P<0.05);Bcl-2蛋白表達明顯下調,差異有統計學意義(t=7.86,P<0.05)。與B組比較,D組Müller細胞Nrf2、HO-1、Bax蛋白表達均下調(t=15.11、26.59、6.27),差異有統計學意義(P<0.05); Bcl-2 蛋白表達上調,差異有統計學意義(t=?6.53,P<0.05)(圖5)。與A組比較,B組Müller細胞Bcl-2/Bax比值降低,差異有統計學意義(t=4.84,P<0.05);與B組比較,不同濃度CPDT干預組Müller細胞Bcl-2/Bax比值增高,差異有統計學意義(t=?7.813,P<0.05)。
圖5
A、B、D組Müller細胞HO-1、Nrf2、Bcl-2、Bax蛋白表達電泳圖
3 討論
CPDT作為二項酶誘導劑,已被證實可在神經元中通過誘導二項酶表達對神經元細胞起保護作用。在肌萎縮側索硬化、腦卒中領域作為誘導二項酶及其相關抗氧化酶發揮其抗氧化應激作用。
DR發生時Müller細胞微環境發生改變,其周圍糖濃度增加,此時周圍的葡萄糖及其自身中間產物產生大量如氧自由基和H2O2等ROS,這些氧自由基可以作用于線粒體通路,對線粒體造成損傷。氧化應激損傷的反應主要通過Nrf2/HO-1通路實現,其中,HO-1可促進代謝性炎癥反應和胰島素抵抗等,糖尿病的發生常伴隨HO-1的增高[9-13]。糖尿病發生時也常伴隨多種器官組織的凋亡。細胞凋亡的線粒體途徑通過Bcl-2在線粒體膜外發揮作用維持線粒體膜的完整性,相反,Bax則破壞線粒體膜完整性,當二者失去平衡時,Bcl-2對抗Bax作用減弱,從而誘導凋亡的發生。作為二項酶誘導劑CPDT可以明顯減輕高糖對Müller細胞的氧化應激作用,并能有效抑制Müller細胞凋亡[11-14]。本研究結果發現,與B組相比較,C、D、E組Müller細胞Nrf2、HO-1、Bax蛋白表達均下調,Bcl-2蛋白表達量增加,表明二項酶誘導劑可通過Nrf2/HO-1通路保護Müller細胞,并抑制細胞的凋亡。另有研究表明二項酶誘導劑對2型糖尿病大鼠視網膜損傷也具有一定保護作用[15]。
本研究結果表明,CPDT可能一方面通過激活Nrf2/HO-1氧化應激通路,另一方面增加Bcl-2蛋白表達量,減少線粒體氧自由基量的生成,從而抑制Müller細胞凋亡。隨著基因技術的發展,可利用基因技術對CPDT在高糖條件中的作用及其機制進行深入研究。
讀者?作者?編者 關于本刊財務收支過程中涉及到的財務管理規定及注意事項說明 本刊財務收支均由承辦單位四川大學華西醫院統一管理。華西醫院賬戶眾多收款項目中,期刊收支項目占比較小,安排的工作人員有限,但管理規定及執行程序同樣非常嚴格。為了提高效率,方便工作,現將版面費等支付過程中涉及到的財務管理規定及注意事項說明如下: 稿件送修并確認刊載后本刊將通知作者按要求數額支付版面費。版面費可通過郵局或銀行支付。 郵局支付者,收款人請寫“編輯部”,不能寄給任何個人。匯款單附言欄必須注明開具發票的抬頭和刊用稿件的編號。發票抬頭若開具單位名稱,應以單位組織機構代碼中的規范名稱為準,如四川大學華西醫院。 通過銀行匯款的版面費均匯至四川大學華西醫院銀行賬戶。匯款者若為個人,應在匯款后當天將匯款憑證掃描或照相后發到編輯部(Email:coretina@163.com);若通過單位對公賬戶匯款,請在提交匯款申請后當日將匯款申請信息告之編輯部,1周內將匯款憑證掃描或照相后發到編輯部。以便及時查找確認并開具發票。若未能按要求時間完成上述工作,發票開具時間則不能確定。跨年的轉款項目財務部門原則上不再查賬并開具發票。 發票出具時,匯款者若為個人,發票抬頭只能開具匯款者姓名;發票抬頭若需開具單位名稱,則只能通過發票抬頭單位對公賬戶匯款。凡依據付款方要求出具一次發票后,不再重新開具任何需要修改發票抬頭等內容的發票。收到版面費后3個月內,以快遞方式寄出版面費發票。 版面費收取及發票查詢的聯系方式,聯系人:萬苗,電話:028-85422060。 除版面費外,其他與本刊相關的財務收支及發票出具注意事項參照上述規定和要求執行。 關于作者與編輯部之間通訊聯系的說明 作者投稿均請通過本刊網站完成,本刊不接受電子郵件或郵寄稿件等其他方式投稿。查詢稿件處理進程、了解說明稿件情況等稿件處理相關的所有溝通聯系均請通過編輯部郵箱coretina@163.com進行。投遞給編輯部的各類信函、匯款均應署名“中華眼底病雜志編輯部”收。上述與工作相關的郵件、信函、匯款均不宜發送、投寄給個人,為便于留痕備查。 編輯部稿件處理流程中相關的所有事宜均同時給第一作者和通信作者聯系,重要事宜應由通信作者承擔責任并負責回復。雜志出版后當期雜志、文章單行本以及稿酬、發票等均只郵寄給第一作者;其第一作者投稿后各種聯系方式有變更者,應及時告知編輯部,以免造成相關工作延誤或不可彌補的損失。 本刊編輯部