引用本文: 漆晨, 東莉潔, 樂毅, 張曉敏, 李筱榮, 郭如如, 茹玉莎, 蘇暢. 多聚嘧啶序列結合蛋白相關剪接因子對體外培養的視網膜色素上皮細胞磷脂酰肌醇3激酶/絲氨酸-蘇氨酸蛋白激酶信號通路的調控作用. 中華眼底病雜志, 2015, 31(4): 363-367. doi: 10.3760/cma.j.issn.1005-1015.2015.04.013 復制
胰島素樣生長因子(IGF-1)是一類促進生長發育和調節物質代謝的多功能活性肽,其介導的血管內皮生長因子(VEGF)表達在視網膜色素上皮(RPE)細胞增生中發揮重要作用[1-3]。而RPE細胞中過表達的VEGF可導致轉基因鼠脈絡膜新生血管的發生[4, 5]。多聚嘧啶序列結合蛋白相關剪接因子(PSF)是存在于細胞核內的多功能蛋白,在IGF-1信號轉導的過程中發揮轉錄抑制因子的作用[6, 7]。玻璃體腔注射重組腺相關病毒包裝的PSF蛋白可抑制氧誘導視網膜病變小鼠視網膜新生血管的形成,下調VEGF的表達[8]。有研究證實,磷脂酰肌醇3 激酶(PI3K)/絲氨酸-蘇氨酸蛋白激酶(Akt)途徑可介導高糖環境中RPE細胞外基質的表達,參與RPE細胞氧化應激反應誘導的凋亡調控[9, 10]。IGF-1可能是通過活化PI3K/Akt信號通路來調控RPE細胞中VEGF的表達[11]。為了進一步明確PSF對RPE細胞IGF-1/VEGF信號通路的影響,我們通過體外實驗觀察了PSF對PI3K/Akt信號通路的調控作用。現將結果報道如下。
1 材料和方法
RPE細胞系ARPE-19細胞由天津醫科大學眼科研究所楊春波教授惠贈。使用含有10%胎牛血清、100 U/ml青霉素和100 U/ml鏈霉素的高糖Dulbecco改良Eagle培養基,置于37℃、95%空氣、5% CO2的密閉恒溫培養箱中培養。參照文獻[12]的方法構建增強型綠色熒光蛋白(pEGFP)-C2-PSF真核表達質粒。pGenesil-PSF-RNAi真核表達質粒購自武漢晶賽公司。PSF單克隆抗體(美國Sigma公司),Akt抗體一抗、磷酸化Akt(pAkt)抗體一抗(美國CST公司)。VEGF:上游引物5′-GGAGGAGGCAT TCGGAAAGT-3′,下游引物5′-GCACTACCTGGGT CAGCTTCA-3′,擴增片段長度為240堿基對(bp); 磷酸甘油醛脫氫酶(GAPDH):上游引物5′-TGAAAC CCTAGTTCATATCTTCAAACA-3′,下游引物5′-CTCCTTCTGTGACTCTAACTTCTCCAT-3′,擴增片段長度為210 bp。
以細胞密度1.5×104個/孔接種ARPE-19細胞于96孔板中,共60孔。取融合密度達70%的細胞用于實驗,分為PSF高表達組、PSF高表達對照組、PSF低表達組、PSF低表達對照組及假轉染組。應用脂質體2000將上調PSF表達的真核質粒pEGFP-C2-PSF、 下調PSF表達的真核質粒pGenesil-PSF-RNAi以0.25、0.50、1.00 μg的遞增量分別轉染至PSF高表達組及PSF低表達組細胞。PSF高表達對照組細胞轉染pEGFP-C2空載質粒。PSF低表達對照組細胞轉染pGenesil-scramble-siRNA質粒。假轉染組僅做轉染處理,不加入任何表達質粒。細胞轉染24 h后更換為無血清培養基,以IGF-1刺激6 h,再次更換為完全培養基繼續培養48 h。采用水溶性四氮唑法檢測有無IGF-1刺激條件下PSF對RPE細胞增生的影響。
采用實時定量聚合酶鏈反應(PCR)檢測PSF對 IGF-1誘導的RPE細胞內VEGF mRNA表達的影響。以細胞密度7.5×105個/孔接種ARPE-19細胞于6孔板中,共6孔。待細胞融合密度達70%時,應用脂質體2000將0.50 μg真核質粒Pegfp-C2-PSF、1.00 μg真核質粒pGenesil-PSF-RNAi及pEGFP-C2空載質粒轉染細胞分別作為PSF高表達組、PSF低表達組、對照組。細胞轉染24 h后更換為無血清培養基,并以IGF-1刺激6 h。提取各組細胞總RNA,逆轉錄為cDNA作為模板,以GAPDH作為內參照。采用2-ΔΔct法分析結果。
采用蛋白免疫印跡法(Western blot)檢測PSF對IGF-1誘導的pAkt、Akt蛋白表達的影響。以細胞密度7.5×105個/孔接種ARPE-19細胞于6孔板中,共8孔。待細胞融合密度達70%時,4孔轉染0.50 μg真核質粒pEGFP-C2-PSF作為PSF高表達組,另4孔只做轉染處理但不加入任何質粒載體作為PSF生理表達組。轉染24 h后,以IGF-1刺激0.5、1.5、3.0 h。收集全細胞提取物,以十二烷基硫酸鈉聚丙烯酰胺凝膠電泳分離蛋白復合物,經半干轉膜后,用5%脫脂奶粉室溫封閉1 h,加入一抗pAkt抗體,室溫孵育2 h,以1倍洗膜緩沖液(TBST)洗膜3次,10 min/次,用辣根過氧化物酶標記的二抗繼續孵育1 h,再以1倍TBST洗膜3次,10 min/次,最后加入增強化學發光曝光底物,進行曝光并拍照。同一張膜經蛋白剝脫處理后,再次以抗Akt抗體作為一抗,進行免疫雜交反應。應用Quantity One軟件對pAkt、Akt灰度值進行分析。
以細胞密度7.5×105個/孔接種ARPE-19細胞于6孔板中,共4孔。第1孔只做轉染處理但不加入任何質粒載體設為對照組;第2孔轉染0.50 μg真核質粒Pegfp-C2-PSF設為單純PSF高表達組;第3孔只做轉染和渥曼青霉素(Wortmannin)處理,但不加入任何質粒載體設為單純渥曼青霉素處理組;第4孔轉染0.50 μg真核質粒pEGFP-C2-PSF并聯合渥曼青霉素處理設為PSF高表達聯合渥曼青霉素處理組。4組細胞均經IGF-1刺激處理6 h后,分別提取各組細胞總RNA,逆轉錄為cDNA作為模板。采用實時定量PCR檢測各組VEGF的表達。
采用SPSS 19.0統計軟件行統計學分析,GraphPad Prism軟件對圖表進行處理。組間比較采用兩因素方差分析及最小顯著差法-t檢驗。P<0.05為差異有統計學意義。
2 結果
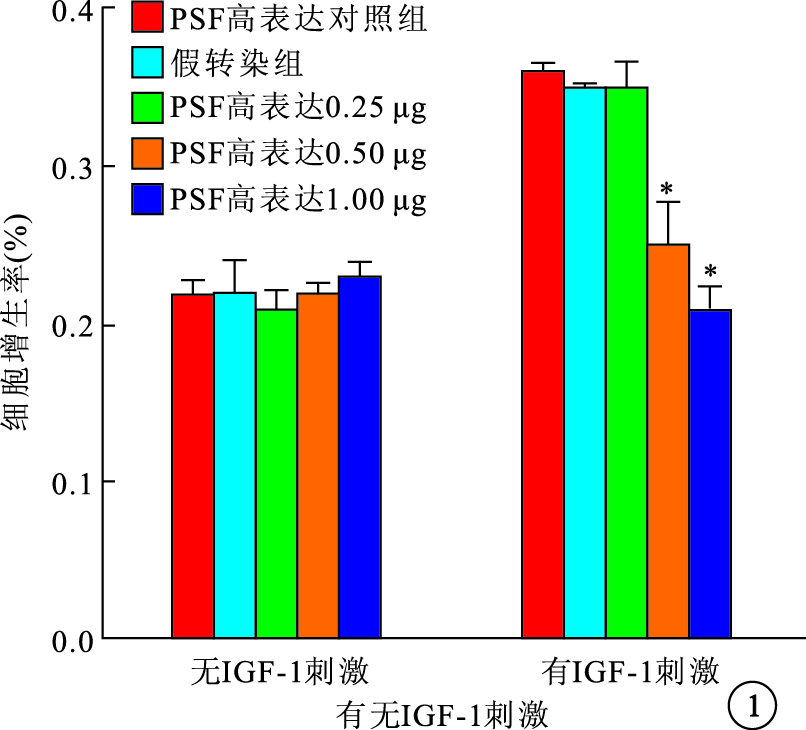
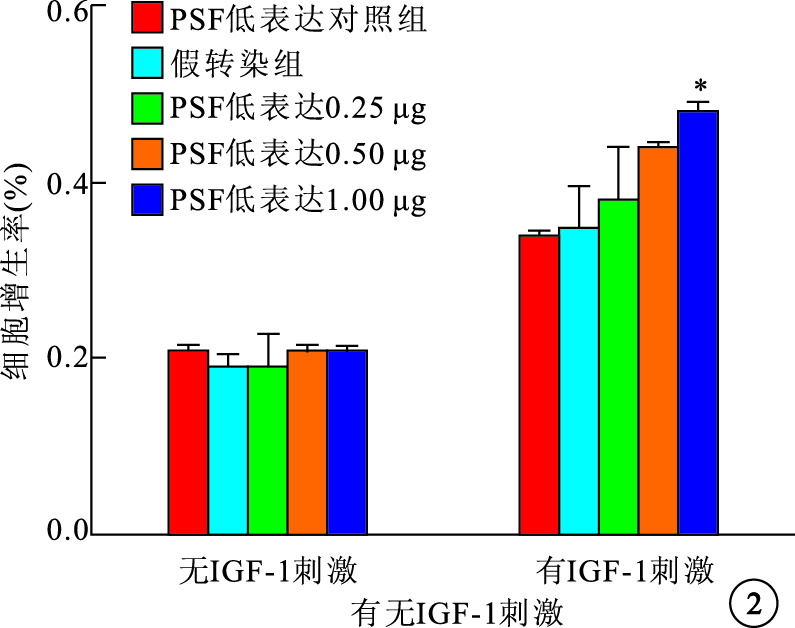
無IGF-1刺激時,PSF高表達和低表達均對RPE細胞的自然增生無影響。IGF-1刺激后,PSF高表達組、PSF高表達對照組及假轉染組之間RPE細胞增生率比較,差異有統計學意義(F=29.728,P<0.05);PSF低表達組、PSF低表達對照組及假轉染組之間RPE細胞增生率比較,差異有統計學意義(F=14.121,P<0.05)。PSF高表達組0.50、1.00 μg真核質粒pEGFP-C2-PSF轉染細胞的增生率較假轉染組細胞增生率明顯降低,差異有統計學意義(P=0.019 1、 0.005 9)(圖 1)。PSF低表達組1.00 μg真核質粒pGenesil-PSF-RNAi轉染細胞的增生率較假轉染組細胞增生率明顯提高,差異有統計學意義(P=0.027 7)(圖 2)。
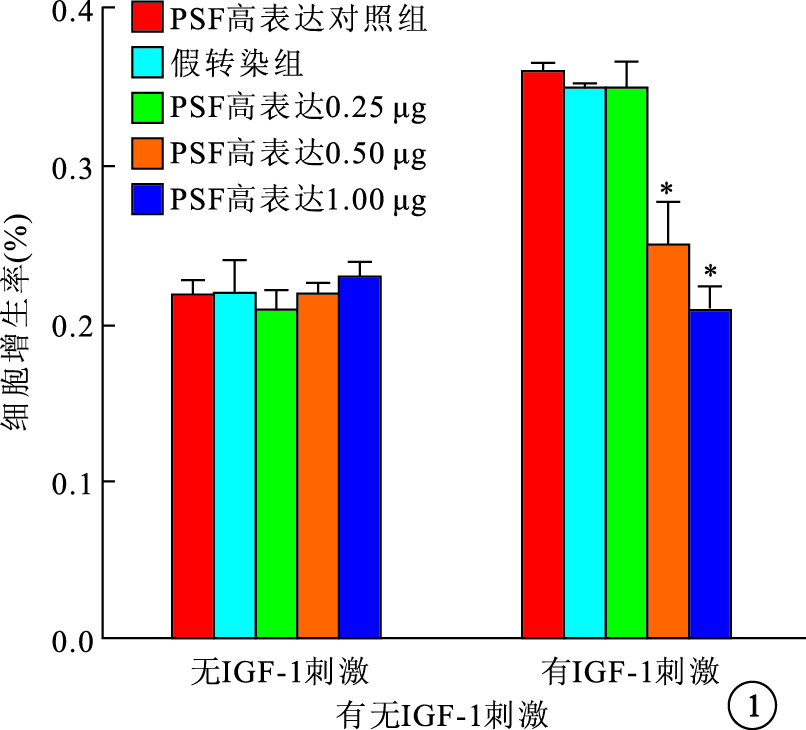 圖1
有無IGF-1刺激時PSF高表達組RPE細胞增生率。*與假轉染組比較,P<0.05
圖1
有無IGF-1刺激時PSF高表達組RPE細胞增生率。*與假轉染組比較,P<0.05
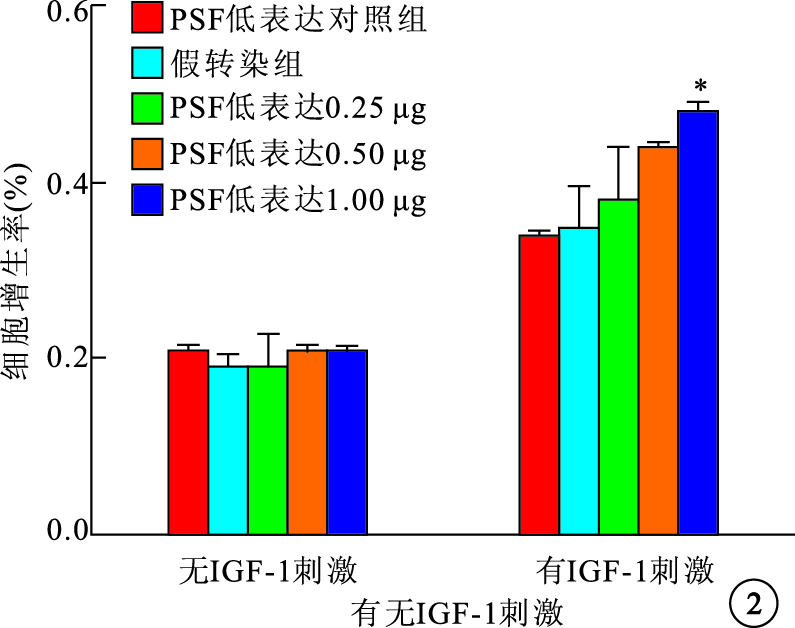 圖2
有無IGF-1刺激時PSF低表達組RPE細胞增生率。*與假轉染組比較,P<0.05
圖2
有無IGF-1刺激時PSF低表達組RPE細胞增生率。*與假轉染組比較,P<0.05
無IGF-1刺激時,PSF高表達和低表達均不影響RPE細胞中VEGF mRNA表達。IGF-1刺激后,PSF高表達組、PSF低表達組及對照組RPE細胞中VEGF mRNA表達分別為0.62±0.06、2.39±0.14、1.81±0.04。與對照組比較,PSF高表達組RPE細胞中VEGF mRNA表達明顯降低,差異有統計學意義(P=0.000 3);PSF低表達組RPE細胞中VEGF mRNA表達明顯提高,差異有統計學意義(P=0.030 9)。
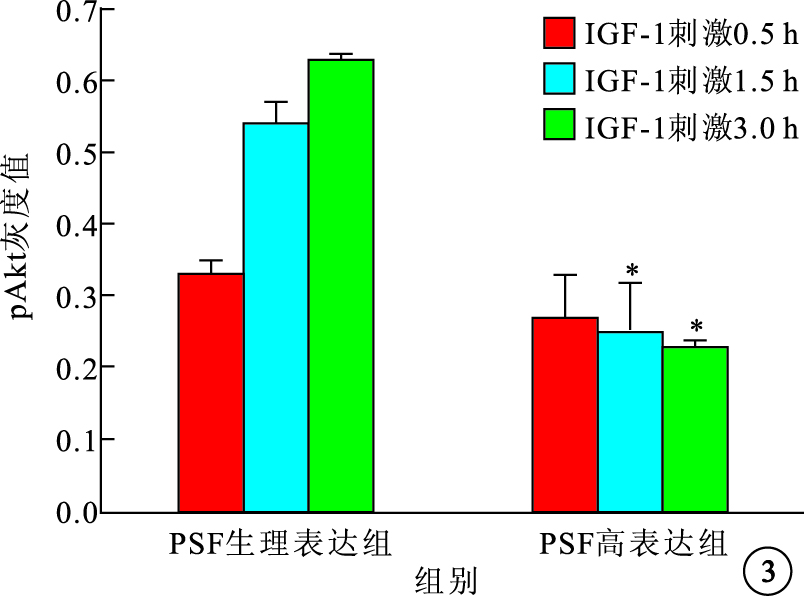
Western blot檢測結果顯示,IGF-1刺激可上調pAkt蛋白表達水平,PSF高表達可明顯下調pAkt蛋白表達水平(圖 3)。與PSF生理表達組比較,IGF-1刺激0.5 h時,PSF高表達組pAkt蛋白表達下調不明顯,差異無統計學意義(P>0.05);IGF-1刺激1.5 h時,PSF高表達組下調pAkt蛋白表達約50%,差異有統計學意義(P=0.022 5);IGF-1刺激3.0 h時,PSF高表達組下調pAkt蛋白表達約65%,差異有統計學意義(P=0.001 3)。各組之間Akt總蛋白表達比較,差異無統計學意義(P>0.05)。
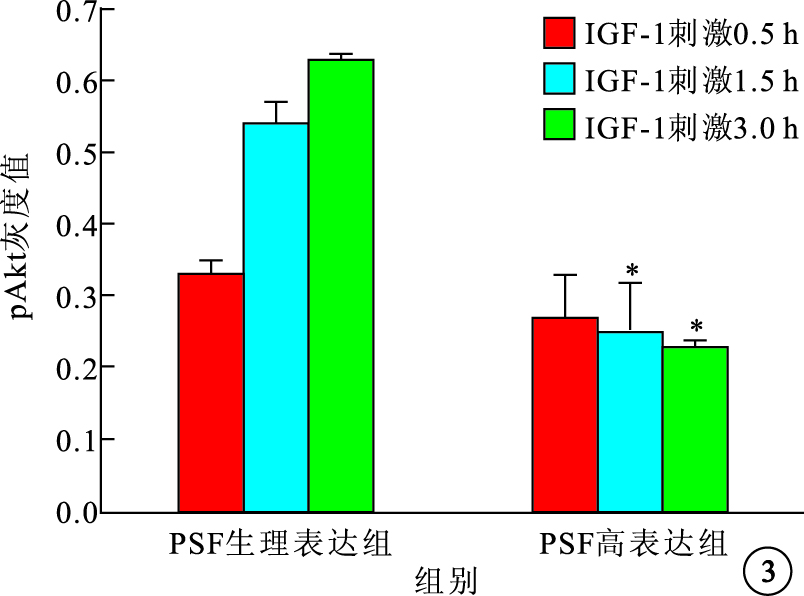 圖3
IGF-1刺激不同時間PSF高表達組及PSF生理表達組pAkt蛋白表達比較。*P<0.05
圖3
IGF-1刺激不同時間PSF高表達組及PSF生理表達組pAkt蛋白表達比較。*P<0.05
IGF-1刺激后,對照組、單純渥曼青霉素處理組、單純PSF高表達組、PSF高表達聯合渥曼青霉素處理組VEGF相對表達分別為2.04±0.10、1.44±0.05、1.53±0.04、0.80±0.04。與對照組比較,單純渥曼青霉素處理組、單純PSF高表達組、PSF高表達聯合渥曼青霉素處理組VEGF相對表達明顯降低,差異均有統計學意義(P=0.004 2、0.004 8、0.002 0)。 與單純渥曼青霉素處理組比較,PSF高表達聯合渥曼青霉素處理組VEGF相對表達明顯降低,差異有統計學意義(P=0.008 6)。
3 討論
Lukong等[12]研究發現,只有在表皮生長因子刺激后,PSF才可通過與BReast腫瘤激酶相結合,參與細胞周期調控。我們的前期研究也證實,IGF-1刺激后,PSF蛋白可抑制視網膜血管內皮細胞中細胞外調節蛋白激酶的活化,從而調控VEGF的表達[13]。可見細胞因子與其受體的結合是PSF發揮作用的啟動信號。PSF通過全身或局部細胞因子水平的改變而感知機體或微環境的動態變化,并做出精準且有針對性的應變反應,來維持內環境的穩態。本研究結果顯示,IGF-1刺激后,PSF高表達可抑制RPE細胞增生,下調pAkt和VEGF的表達;但在無IGF-1刺激時,PSF的上述作用均不能發揮。說明只有在IGF-1刺激的前提下,高表達PSF才能發揮對RPE細胞中VEGF表達的抑制作用。提示PSF對于RPE細胞的調控作用具有明顯的IGF-1配體依賴性。這一發現有著重要的臨床意義。因為生理水平的VEGF可以維持血管的正常結構和功能,過度抑制對正常血管的功能有所影響,引起一系列的遠期并發癥。所以,臨床中抗VEGF治療的理想狀態是可以選擇性的抑制病理性升高的VEGF。PSF通過感知眼部微環境中IGF-1水平的改變,準確的識別VEGF病理性升高及其生理水平,既能有效抑制病理性升高的VEGF,又不影響其生理水平的維持。提示PSF有望成為特異性抑制病理性VEGF表達的新靶點。
鑒于PSF高表達之后RPE細胞中pAkt蛋白表達水平呈下調趨勢,為進一步探討其作用機制,我們將Akt信號通路的特異性抑制劑渥曼青霉素引入到實驗體系中,以證明PSF是否通過影響PI3K/Akt 通路的活性來發揮作用。Weng等[14]研究發現,IGF-1可以通過活化RPE細胞中絲裂原活化蛋白激酶(MAPK)途徑來上調VEGF表達,而MAPK途徑特異性抑制劑PD98059可顯著拮抗IGF-1的這一作用。說明IGF-1是通過活化MAPK途徑來發揮其作用。渥曼青霉素是一種常用的PI3K抑制劑,可以通透細胞,并與PI3K的催化亞基相結合,特異性抑制PI3K,從而抑制PI3K/Akt信號途徑[15, 16]。本研究結果顯示,IGF-1刺激后將高表達的PSF與渥曼青霉素聯合應用處理細胞時,其對于VEGF表達的下降調節作用顯著高于任何一種單獨處理情況。說明高表達PSF與渥曼青霉素在抑制VEGF表達方面可發揮累加作用。這也從另一個角度證明了PSF在一定程度上是通過抑制IGF-1刺激后Akt信號途徑的活化來下調VEGF的表達,PSF可以負性調控IGF-1/PI3K/Akt信號通路的活性。
本研究結果表明,PSF能抑制IGF-1 刺激后體外培養的RPE細胞PI3K/Akt信號通路活化,下調VEGF表達。但由于本研究只是探討分析了PSF對于RPE細胞中VEGF表達的調控作用及可能機制,未針對PSF對RPE細胞遷移、吞噬、營養影響進行觀察。所以,有關PSF蛋白對RPE細胞生物學功能的調控作用還有待進一步深入研究,從而為PSF在RPE相關性疾病中的作用研究提供實驗依據。
胰島素樣生長因子(IGF-1)是一類促進生長發育和調節物質代謝的多功能活性肽,其介導的血管內皮生長因子(VEGF)表達在視網膜色素上皮(RPE)細胞增生中發揮重要作用[1-3]。而RPE細胞中過表達的VEGF可導致轉基因鼠脈絡膜新生血管的發生[4, 5]。多聚嘧啶序列結合蛋白相關剪接因子(PSF)是存在于細胞核內的多功能蛋白,在IGF-1信號轉導的過程中發揮轉錄抑制因子的作用[6, 7]。玻璃體腔注射重組腺相關病毒包裝的PSF蛋白可抑制氧誘導視網膜病變小鼠視網膜新生血管的形成,下調VEGF的表達[8]。有研究證實,磷脂酰肌醇3 激酶(PI3K)/絲氨酸-蘇氨酸蛋白激酶(Akt)途徑可介導高糖環境中RPE細胞外基質的表達,參與RPE細胞氧化應激反應誘導的凋亡調控[9, 10]。IGF-1可能是通過活化PI3K/Akt信號通路來調控RPE細胞中VEGF的表達[11]。為了進一步明確PSF對RPE細胞IGF-1/VEGF信號通路的影響,我們通過體外實驗觀察了PSF對PI3K/Akt信號通路的調控作用。現將結果報道如下。
1 材料和方法
RPE細胞系ARPE-19細胞由天津醫科大學眼科研究所楊春波教授惠贈。使用含有10%胎牛血清、100 U/ml青霉素和100 U/ml鏈霉素的高糖Dulbecco改良Eagle培養基,置于37℃、95%空氣、5% CO2的密閉恒溫培養箱中培養。參照文獻[12]的方法構建增強型綠色熒光蛋白(pEGFP)-C2-PSF真核表達質粒。pGenesil-PSF-RNAi真核表達質粒購自武漢晶賽公司。PSF單克隆抗體(美國Sigma公司),Akt抗體一抗、磷酸化Akt(pAkt)抗體一抗(美國CST公司)。VEGF:上游引物5′-GGAGGAGGCAT TCGGAAAGT-3′,下游引物5′-GCACTACCTGGGT CAGCTTCA-3′,擴增片段長度為240堿基對(bp); 磷酸甘油醛脫氫酶(GAPDH):上游引物5′-TGAAAC CCTAGTTCATATCTTCAAACA-3′,下游引物5′-CTCCTTCTGTGACTCTAACTTCTCCAT-3′,擴增片段長度為210 bp。
以細胞密度1.5×104個/孔接種ARPE-19細胞于96孔板中,共60孔。取融合密度達70%的細胞用于實驗,分為PSF高表達組、PSF高表達對照組、PSF低表達組、PSF低表達對照組及假轉染組。應用脂質體2000將上調PSF表達的真核質粒pEGFP-C2-PSF、 下調PSF表達的真核質粒pGenesil-PSF-RNAi以0.25、0.50、1.00 μg的遞增量分別轉染至PSF高表達組及PSF低表達組細胞。PSF高表達對照組細胞轉染pEGFP-C2空載質粒。PSF低表達對照組細胞轉染pGenesil-scramble-siRNA質粒。假轉染組僅做轉染處理,不加入任何表達質粒。細胞轉染24 h后更換為無血清培養基,以IGF-1刺激6 h,再次更換為完全培養基繼續培養48 h。采用水溶性四氮唑法檢測有無IGF-1刺激條件下PSF對RPE細胞增生的影響。
采用實時定量聚合酶鏈反應(PCR)檢測PSF對 IGF-1誘導的RPE細胞內VEGF mRNA表達的影響。以細胞密度7.5×105個/孔接種ARPE-19細胞于6孔板中,共6孔。待細胞融合密度達70%時,應用脂質體2000將0.50 μg真核質粒Pegfp-C2-PSF、1.00 μg真核質粒pGenesil-PSF-RNAi及pEGFP-C2空載質粒轉染細胞分別作為PSF高表達組、PSF低表達組、對照組。細胞轉染24 h后更換為無血清培養基,并以IGF-1刺激6 h。提取各組細胞總RNA,逆轉錄為cDNA作為模板,以GAPDH作為內參照。采用2-ΔΔct法分析結果。
采用蛋白免疫印跡法(Western blot)檢測PSF對IGF-1誘導的pAkt、Akt蛋白表達的影響。以細胞密度7.5×105個/孔接種ARPE-19細胞于6孔板中,共8孔。待細胞融合密度達70%時,4孔轉染0.50 μg真核質粒pEGFP-C2-PSF作為PSF高表達組,另4孔只做轉染處理但不加入任何質粒載體作為PSF生理表達組。轉染24 h后,以IGF-1刺激0.5、1.5、3.0 h。收集全細胞提取物,以十二烷基硫酸鈉聚丙烯酰胺凝膠電泳分離蛋白復合物,經半干轉膜后,用5%脫脂奶粉室溫封閉1 h,加入一抗pAkt抗體,室溫孵育2 h,以1倍洗膜緩沖液(TBST)洗膜3次,10 min/次,用辣根過氧化物酶標記的二抗繼續孵育1 h,再以1倍TBST洗膜3次,10 min/次,最后加入增強化學發光曝光底物,進行曝光并拍照。同一張膜經蛋白剝脫處理后,再次以抗Akt抗體作為一抗,進行免疫雜交反應。應用Quantity One軟件對pAkt、Akt灰度值進行分析。
以細胞密度7.5×105個/孔接種ARPE-19細胞于6孔板中,共4孔。第1孔只做轉染處理但不加入任何質粒載體設為對照組;第2孔轉染0.50 μg真核質粒Pegfp-C2-PSF設為單純PSF高表達組;第3孔只做轉染和渥曼青霉素(Wortmannin)處理,但不加入任何質粒載體設為單純渥曼青霉素處理組;第4孔轉染0.50 μg真核質粒pEGFP-C2-PSF并聯合渥曼青霉素處理設為PSF高表達聯合渥曼青霉素處理組。4組細胞均經IGF-1刺激處理6 h后,分別提取各組細胞總RNA,逆轉錄為cDNA作為模板。采用實時定量PCR檢測各組VEGF的表達。
采用SPSS 19.0統計軟件行統計學分析,GraphPad Prism軟件對圖表進行處理。組間比較采用兩因素方差分析及最小顯著差法-t檢驗。P<0.05為差異有統計學意義。
2 結果
無IGF-1刺激時,PSF高表達和低表達均對RPE細胞的自然增生無影響。IGF-1刺激后,PSF高表達組、PSF高表達對照組及假轉染組之間RPE細胞增生率比較,差異有統計學意義(F=29.728,P<0.05);PSF低表達組、PSF低表達對照組及假轉染組之間RPE細胞增生率比較,差異有統計學意義(F=14.121,P<0.05)。PSF高表達組0.50、1.00 μg真核質粒pEGFP-C2-PSF轉染細胞的增生率較假轉染組細胞增生率明顯降低,差異有統計學意義(P=0.019 1、 0.005 9)(圖 1)。PSF低表達組1.00 μg真核質粒pGenesil-PSF-RNAi轉染細胞的增生率較假轉染組細胞增生率明顯提高,差異有統計學意義(P=0.027 7)(圖 2)。
圖1
有無IGF-1刺激時PSF高表達組RPE細胞增生率。*與假轉染組比較,P<0.05
圖2
有無IGF-1刺激時PSF低表達組RPE細胞增生率。*與假轉染組比較,P<0.05
無IGF-1刺激時,PSF高表達和低表達均不影響RPE細胞中VEGF mRNA表達。IGF-1刺激后,PSF高表達組、PSF低表達組及對照組RPE細胞中VEGF mRNA表達分別為0.62±0.06、2.39±0.14、1.81±0.04。與對照組比較,PSF高表達組RPE細胞中VEGF mRNA表達明顯降低,差異有統計學意義(P=0.000 3);PSF低表達組RPE細胞中VEGF mRNA表達明顯提高,差異有統計學意義(P=0.030 9)。
Western blot檢測結果顯示,IGF-1刺激可上調pAkt蛋白表達水平,PSF高表達可明顯下調pAkt蛋白表達水平(圖 3)。與PSF生理表達組比較,IGF-1刺激0.5 h時,PSF高表達組pAkt蛋白表達下調不明顯,差異無統計學意義(P>0.05);IGF-1刺激1.5 h時,PSF高表達組下調pAkt蛋白表達約50%,差異有統計學意義(P=0.022 5);IGF-1刺激3.0 h時,PSF高表達組下調pAkt蛋白表達約65%,差異有統計學意義(P=0.001 3)。各組之間Akt總蛋白表達比較,差異無統計學意義(P>0.05)。
圖3
IGF-1刺激不同時間PSF高表達組及PSF生理表達組pAkt蛋白表達比較。*P<0.05
IGF-1刺激后,對照組、單純渥曼青霉素處理組、單純PSF高表達組、PSF高表達聯合渥曼青霉素處理組VEGF相對表達分別為2.04±0.10、1.44±0.05、1.53±0.04、0.80±0.04。與對照組比較,單純渥曼青霉素處理組、單純PSF高表達組、PSF高表達聯合渥曼青霉素處理組VEGF相對表達明顯降低,差異均有統計學意義(P=0.004 2、0.004 8、0.002 0)。 與單純渥曼青霉素處理組比較,PSF高表達聯合渥曼青霉素處理組VEGF相對表達明顯降低,差異有統計學意義(P=0.008 6)。
3 討論
Lukong等[12]研究發現,只有在表皮生長因子刺激后,PSF才可通過與BReast腫瘤激酶相結合,參與細胞周期調控。我們的前期研究也證實,IGF-1刺激后,PSF蛋白可抑制視網膜血管內皮細胞中細胞外調節蛋白激酶的活化,從而調控VEGF的表達[13]。可見細胞因子與其受體的結合是PSF發揮作用的啟動信號。PSF通過全身或局部細胞因子水平的改變而感知機體或微環境的動態變化,并做出精準且有針對性的應變反應,來維持內環境的穩態。本研究結果顯示,IGF-1刺激后,PSF高表達可抑制RPE細胞增生,下調pAkt和VEGF的表達;但在無IGF-1刺激時,PSF的上述作用均不能發揮。說明只有在IGF-1刺激的前提下,高表達PSF才能發揮對RPE細胞中VEGF表達的抑制作用。提示PSF對于RPE細胞的調控作用具有明顯的IGF-1配體依賴性。這一發現有著重要的臨床意義。因為生理水平的VEGF可以維持血管的正常結構和功能,過度抑制對正常血管的功能有所影響,引起一系列的遠期并發癥。所以,臨床中抗VEGF治療的理想狀態是可以選擇性的抑制病理性升高的VEGF。PSF通過感知眼部微環境中IGF-1水平的改變,準確的識別VEGF病理性升高及其生理水平,既能有效抑制病理性升高的VEGF,又不影響其生理水平的維持。提示PSF有望成為特異性抑制病理性VEGF表達的新靶點。
鑒于PSF高表達之后RPE細胞中pAkt蛋白表達水平呈下調趨勢,為進一步探討其作用機制,我們將Akt信號通路的特異性抑制劑渥曼青霉素引入到實驗體系中,以證明PSF是否通過影響PI3K/Akt 通路的活性來發揮作用。Weng等[14]研究發現,IGF-1可以通過活化RPE細胞中絲裂原活化蛋白激酶(MAPK)途徑來上調VEGF表達,而MAPK途徑特異性抑制劑PD98059可顯著拮抗IGF-1的這一作用。說明IGF-1是通過活化MAPK途徑來發揮其作用。渥曼青霉素是一種常用的PI3K抑制劑,可以通透細胞,并與PI3K的催化亞基相結合,特異性抑制PI3K,從而抑制PI3K/Akt信號途徑[15, 16]。本研究結果顯示,IGF-1刺激后將高表達的PSF與渥曼青霉素聯合應用處理細胞時,其對于VEGF表達的下降調節作用顯著高于任何一種單獨處理情況。說明高表達PSF與渥曼青霉素在抑制VEGF表達方面可發揮累加作用。這也從另一個角度證明了PSF在一定程度上是通過抑制IGF-1刺激后Akt信號途徑的活化來下調VEGF的表達,PSF可以負性調控IGF-1/PI3K/Akt信號通路的活性。
本研究結果表明,PSF能抑制IGF-1 刺激后體外培養的RPE細胞PI3K/Akt信號通路活化,下調VEGF表達。但由于本研究只是探討分析了PSF對于RPE細胞中VEGF表達的調控作用及可能機制,未針對PSF對RPE細胞遷移、吞噬、營養影響進行觀察。所以,有關PSF蛋白對RPE細胞生物學功能的調控作用還有待進一步深入研究,從而為PSF在RPE相關性疾病中的作用研究提供實驗依據。