引用本文: 韓娟娟, 戴旭鋒, 張華, 戚艷, 龐繼景. 溶血卵磷脂酰基轉移酶1自發突變小鼠視網膜組織和閃光視網膜電圖改變. 中華眼底病雜志, 2014, 30(3): 289-293. doi: 10.3760/cma.j.issn.1005-1015.2014.03.015 復制
溶血卵磷脂酰基轉移酶1(LPCAT1)是一種促進磷脂合成及重構的酶,能使溶血卵磷脂轉化成二棕櫚酰卵磷脂,在人類多處正常組織中存在表達[1, 2]。研究發現,小鼠Lpcat1基因自發突變可引起視網膜變性,小鼠13號染色體上Lpcat1基因第3外顯子由于單個核苷酸插入引起移碼突變是rd11小鼠視網膜致病原因[3, 4]。但目前有關LPCAT1的研究多集中于合成肺表面活性物質及腫瘤[2, 5-10]。而針對其與視網膜的關系研究還較為少見。為此,我們觀察Lpcat1基因自發突變小鼠視網膜組織和閃光視網膜電圖(F-ERG)改變情況,現將結果報道如下。
1 材料和方法
Lpcat1基因自發突變純合子的rd11新生小鼠60只,美國實驗動物中心提供,作為實驗組。與之同齡的野生型C57BL/6J小鼠60只,溫州醫科大學實驗動物中心提供,作為對照組。
小鼠8周齡時,每組取5只小鼠。采用CO2氣體處死小鼠并摘取眼球,置于4%多聚甲醛冰凍切片固定液中4 ℃過夜,將固定好的標本在30%蔗糖溶液中脫水5 h。眼球置于冷凍包埋液中-80 ℃包埋,冰凍切片機做厚度為10 μm連續切片,切片平行于視軸方向,貼片于0.05%多聚賴氨酸處理的載玻片上,室溫干燥1 h。冰凍切片置于0.3%Triton X-100穿透液中30 min增強細胞膜通透性,5%牛血清白蛋白工作液中37 ℃封閉2 h,滴加一抗兔抗小鼠LPCAT1抗體(美國Sigma公司),4 ℃孵育過夜,二抗紅色熒光染料Cy3標記的山羊抗兔IgG(德國Millipore公司)37 ℃孵育1 h,清洗后滴加4,6-聯脒-2-苯基吲哚(DAPI)染色細胞核。熒光顯微鏡下觀察視網膜內核層(INL)、內叢狀層(IPL)、外核層(ONL)、外叢狀層(OPL)、神經節細胞層(GCL)、光感受器內節(IS)及外節(OS)等組織,根據是否產生紅色熒光確定LPCAT1陽性或陰性表達。
出生后3、6、9 d和2、4、6、8周,每組各時間點取5只小鼠。采用CO2氣體處死小鼠并摘取眼球,置于含4%多聚甲醛、2%三氯乙酸、2%氯化鋅和20%異丙醇的石蠟切片固定液中,4 ℃固定24 h;將固定好的標本常規脫水、浸蠟和包埋,將眼球蠟塊平行于視軸方向切片,取經視網膜后極部,距視盤邊緣大約50 μm區域的切片,片厚4 μm,貼片于0.05%多聚賴氨酸處理的載玻片上。常規蘇木精-伊紅(HE)染色后中性樹膠封片,生物顯微鏡下觀察視網膜結構。
出生后2、4、6、8周,每組各時間點取5只小鼠,行單眼全視野F-ERG檢查。小鼠暗適應過夜,在暗紅光條件下用0.5%復方托吡卡胺滴眼液散瞳,分別以72 mg/kg和4 mg/kg的劑量腹腔注射氯胺酮和賽拉嗪混合液麻醉。小鼠全身麻醉后在右眼眼球角鞏膜緣放置由直徑2 mm細銀絲制成的記錄電極,雙耳正中頭皮下和尾部皮下分別放置自制針狀的參考和接地電極。補暗適應5 min后先記錄暗視桿體反應,然后明適應10 min后再記錄明視錐體反應。使用刺激器(Ganzfeld Q450,德國Roland公司),白色閃光的色溫為7000 K。參照文獻[11]的方法設置參數,誘導暗視桿體反應的光強度為0.01 cd·s/m2;明適應背景白光的亮度為30 cd/m2,標準閃光強度3.0 cd·s/m2誘導明視錐體反應;單次閃光刺激持續時間2 ms,刺激間隔10 s,重復5次后取平均值,通頻帶設為1~100 Hz。 選取反應較大的正向b波作為觀察指標。
采用 SPSS 13.0 統計軟件對數據進行統計分析,實驗數據均以均數±標準差(
2 結果
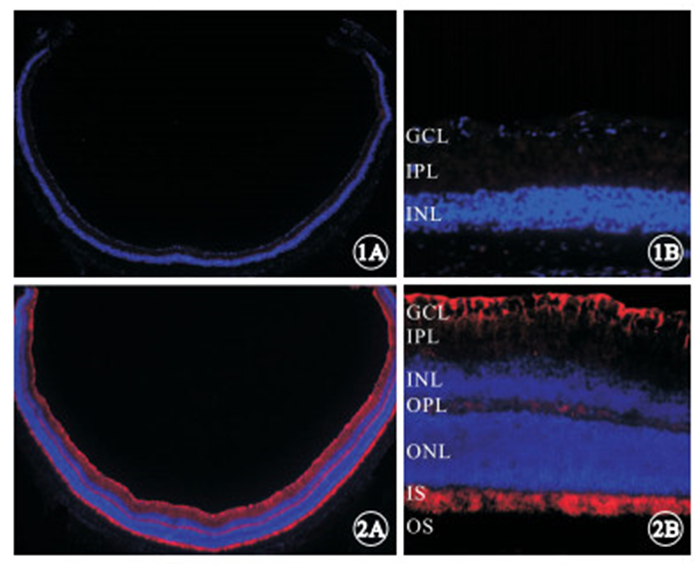
免疫組織化學染色發現,實驗組小鼠視網膜組織中LPCAT1呈陰性表達(圖 1);對照組小鼠視網膜組織中LPCAT1呈陽性表達,主要分布于IS,GCL可見少量表達(圖 2)。
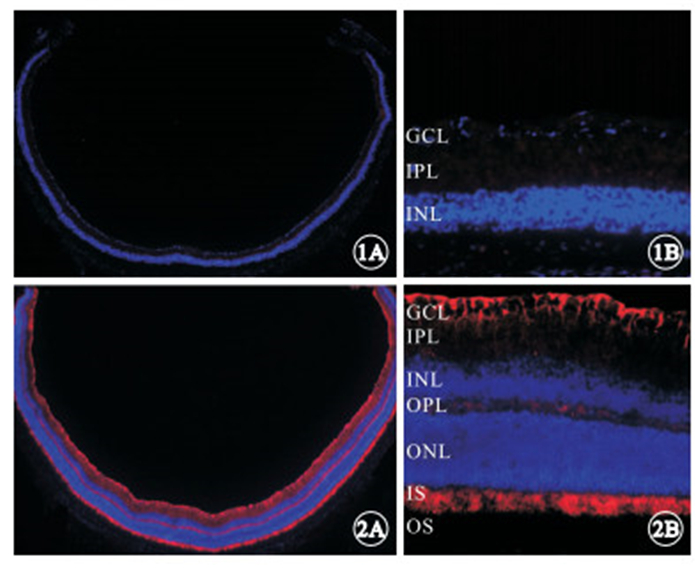 圖1
實驗組小鼠出生后8周LPCAT1表達的熒光顯微鏡像。1A. 視網膜后極部和周邊部均呈陰性表達 免疫熒光染色 ×25;1B. 視網膜后極部距視盤邊緣大約50 μm區域殘存的INL、IPL、GCL各層均呈陰性表達 免疫熒光染色 ×200 ? ?圖 2 對照組小鼠出生后8周LPCAT1表達的熒光顯微鏡像。2A. 視網膜后極部和周邊部均呈陽性表達 免疫熒光染色 ×25;2B. 視網膜后極部距視盤邊緣大約50 μm區域IS可見高密度表達,GCL見較少量表達 免疫熒光染色 ×200
圖1
實驗組小鼠出生后8周LPCAT1表達的熒光顯微鏡像。1A. 視網膜后極部和周邊部均呈陰性表達 免疫熒光染色 ×25;1B. 視網膜后極部距視盤邊緣大約50 μm區域殘存的INL、IPL、GCL各層均呈陰性表達 免疫熒光染色 ×200 ? ?圖 2 對照組小鼠出生后8周LPCAT1表達的熒光顯微鏡像。2A. 視網膜后極部和周邊部均呈陽性表達 免疫熒光染色 ×25;2B. 視網膜后極部距視盤邊緣大約50 μm區域IS可見高密度表達,GCL見較少量表達 免疫熒光染色 ×200
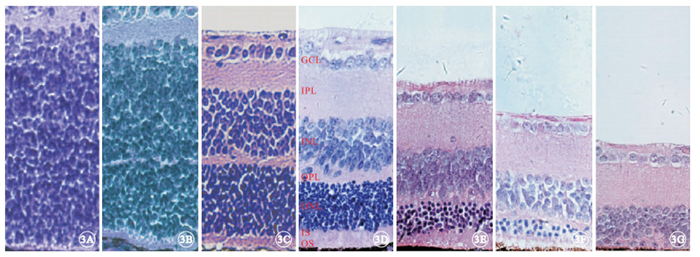
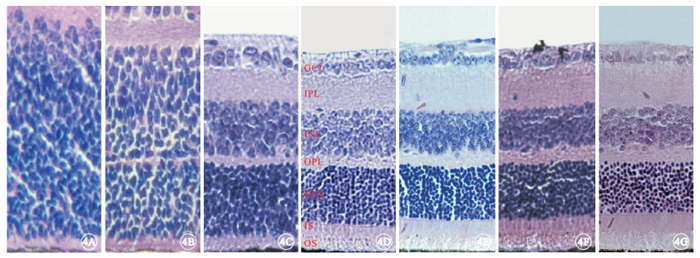
光學顯微鏡觀察發現,實驗組小鼠于出生后9 d左右視網膜ONL已基本形成(圖 3A~3C);出生后2周ONL有8~9層結構(圖 3D);出生后4周ONL減少至5~6層結構(圖 3E),光感受器內外節連接(IS/OS)厚度變窄,并可見一些細胞核殘骸向視網膜色素上皮(RPE)端靠近,OPL厚度也相應變窄;出生后6周ONL僅剩2層結構(圖 3F);出生后8周ONL殘留0~1層結構(圖 3G)。對照組小鼠視網膜組織中ONL也在出生后9 d左右形成(圖 4A~4C);出生后2~8周,ONL始終保持在11~12層結構(圖 4D~4G),且IS/OS及OPL厚度均基本正常。
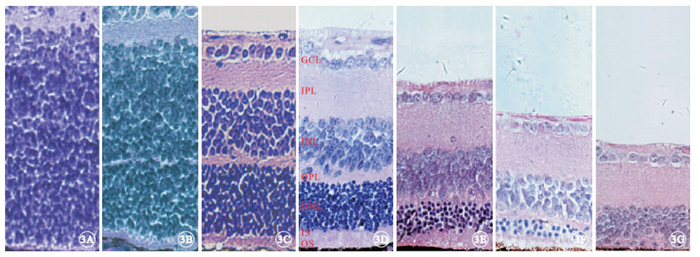 圖3
實驗組小鼠視網膜組織結構光學顯微鏡像。3A~3G分別為出生后3、6、9 d和2、4、6、8周。出生后9 d左右視網膜ONL已基本形成,出生后4~8周ONL層數迅速減少,直至幾乎消失,IS/OS及OPL厚度均變窄 HE ×400
圖3
實驗組小鼠視網膜組織結構光學顯微鏡像。3A~3G分別為出生后3、6、9 d和2、4、6、8周。出生后9 d左右視網膜ONL已基本形成,出生后4~8周ONL層數迅速減少,直至幾乎消失,IS/OS及OPL厚度均變窄 HE ×400
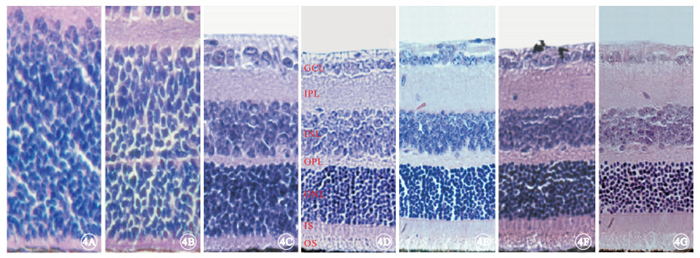 圖4
對照組小鼠視網膜組織結構光學顯微鏡像。4A~4G分別為出生后3、6、9 d和2、4、6、8周。出生后9 d左右視網膜ONL已基本形成,出生后2~8周時視網膜各層組織結構正常 HE ×400
圖4
對照組小鼠視網膜組織結構光學顯微鏡像。4A~4G分別為出生后3、6、9 d和2、4、6、8周。出生后9 d左右視網膜ONL已基本形成,出生后2~8周時視網膜各層組織結構正常 HE ×400
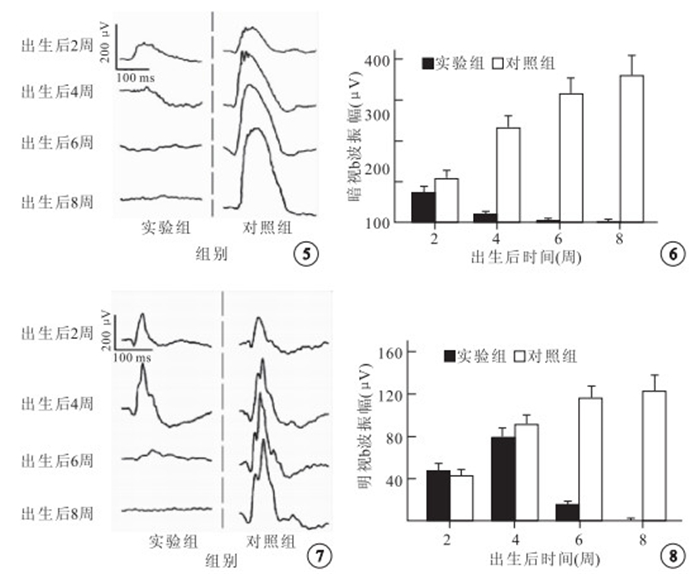
全視野F-ERG檢測結果顯示,實驗組小鼠出生后2、4周暗視桿體系統反應存在,呈降低趨勢,出生后6~8周暗視桿體系統反應消失;對照組小鼠各時間點暗視桿體系統反應正常(圖 5)。 出生后2周,實驗組和對照組小鼠b波振幅分別為(72.8±15.6)、(105.2± 21.1) μV,實驗組較對照組降低,但差異無統計學意義(t=-2.760,P=0.025)。出生后4周,實驗組和對照組小鼠b波振幅分別為(20.6±6.4)、(231.8±32.0) μV,實驗組較對照組明顯降低,差異有統計學意義(t=-14.471,P=0.000)(圖 6)。
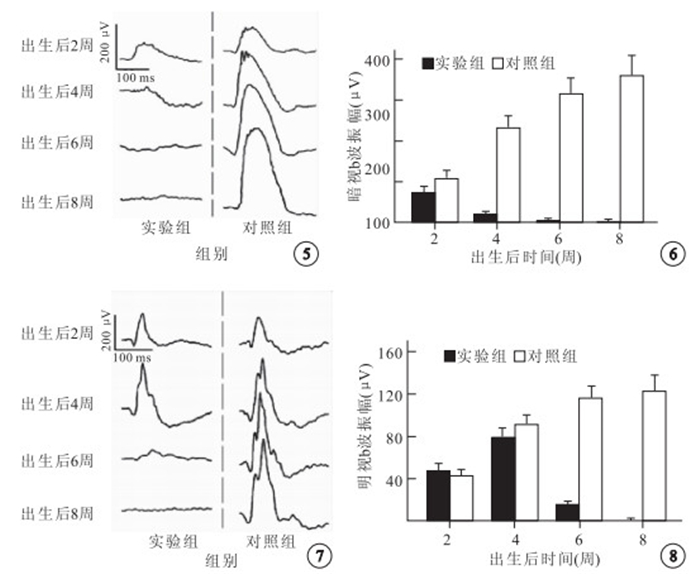 圖5
兩組小鼠暗適應狀態下全視野F-ERG檢測像。實驗組小鼠出生后2、4周桿體系統反應存在,但反應幅值較同齡對照降低,出生后6~8周桿體系統反應消失;對照組小鼠各時間點暗視桿體系統反應正常 ? ?圖 6 兩組小鼠暗視b波振幅比較 ? ?圖 7 兩組小鼠明適應狀態下全視野F-ERG檢測像。實驗組小鼠出生后2、4、6周錐體系統反應存在,但反應幅值隨年齡增長逐漸降低,出生后8周明視錐體系統反應消失;對照組小鼠各時間點明視錐體系統反應正常 ? ?圖 8 兩組小鼠明視b波振幅比較
圖5
兩組小鼠暗適應狀態下全視野F-ERG檢測像。實驗組小鼠出生后2、4周桿體系統反應存在,但反應幅值較同齡對照降低,出生后6~8周桿體系統反應消失;對照組小鼠各時間點暗視桿體系統反應正常 ? ?圖 6 兩組小鼠暗視b波振幅比較 ? ?圖 7 兩組小鼠明適應狀態下全視野F-ERG檢測像。實驗組小鼠出生后2、4、6周錐體系統反應存在,但反應幅值隨年齡增長逐漸降低,出生后8周明視錐體系統反應消失;對照組小鼠各時間點明視錐體系統反應正常 ? ?圖 8 兩組小鼠明視b波振幅比較
全視野F-ERG檢測結果顯示,實驗組小鼠出生后2、4、6周明視錐體系統反應存在,呈降低趨勢,出生后8周明視錐體系統反應消失;對照組小鼠各時間點明視錐體系統反應正常(圖 7)。出生后2、4周,實驗組小鼠b波振幅分別為(46.8±7.2)、(78.0±8.2) μV;對照組小鼠b波振幅分別為(42.8±6.4)、(91.4±9.4) μV。兩組比較,差異均無統計學意義(t=0.930、-2.401,P=0.379、0.043)。出生后6周,實驗組和對照組小鼠b波振幅分別為(17.2±2.0)、(116.2±12.9) μV,實驗組較對照組明顯降低,差異有統計學意義(t=-17.008,P=0.000)(圖 8)。
3 討論
LPCAT1的相對分子質量為59×103,位于內質網,其功能為促進脂質合成和代謝,并影響Ca2+通道[12]。有研究發現,糖尿病小鼠視網膜的LPCAT1表達降低[13]。Friedman等[3]研究發現,Lpcat1基因突變與小鼠視網膜變性有關,他們用定量聚合酶鏈反應的方法在野生型小鼠視網膜組織的INL、ONL及GCL均檢測到Lpcat1基因表達。我們采用免疫熒光染色法檢測LPCAT1在全視網膜的表達情況,結果顯示實驗組rd11小鼠全視網膜組織LPCAT1呈陰性表達。而對照組小鼠視網膜組織中LPCAT1呈陽性表達,其中IS可見高密度表達,GCL較少表達,但在INL呈陰性表達,這可能與其表達量太低有關[3]。這一結果與LPCAT1位于內質網,而內質網在光感受器中的分布主要集中在IS的結果相吻合[12, 14]。
LPCAT1缺陷引起視網膜變性的詳細過程目前仍不太清楚。本研究選取小鼠出生后3、6、9 d和2、4、6、8周7個時間點,對視網膜組織顯微結構進行動態觀察。結果顯示,實驗組rd11小鼠視網膜光感受器細胞在出生后9 d左右形成,從時間上判斷與對照組野生型小鼠差別不大。說明LPCAT1缺陷并未引起視網膜組織發育的明顯滯后。從小鼠各年齡段視網膜組織形態分析,LPCAT1缺陷主要引起光感受器細胞快速喪失,這可能與細胞凋亡有關。已有研究發現,LPCAT1缺陷可導致Ca2+通道持續開放,細胞內Ca2+濃度過高;鈣超載后引起一系列生化反應,最終導致細胞凋亡通路的啟動[14, 15]。而由于LPCAT1在正常視網膜組織中的GCL較少表達,該蛋白缺陷對后者結構的影響可能不大,所以我們未發現GCL形態的明顯改變。
F-ERG是視網膜組織接受光刺激后產生的一組電位反應[16]。小鼠完全暗適應狀態下,用弱白光刺激,可以引出桿體系統反應;經過明適應,充分抑制桿體系統功能后,用標準閃光強度可以誘導出錐體系統反應[11]。本研究結果顯示,對照組野生型小鼠出生后2~6周,其視網膜功能尚未完全發育成熟,直至出生后8周F-ERG才基本穩定。我們發現,實驗組rd11小鼠視桿細胞變性始于出生后4周,視錐細胞變性始于出生后6周,在接下去不到2周的時間內視網膜功能幾乎完全喪失。而視網膜結構也顯示小鼠出生后4周光感受器減少最為顯著,這與在數量上占絕對優勢的視桿細胞的變性凋亡基本相符;出生后6~8周,視網膜殘留的具有椎體功能的1~2層光感受器也完全消失了。這些結果提示光感受器變性發生時間早,進展速度快。另外,視桿細胞變性要早于視錐細胞,這同臨床上原發性視網膜色素變性發病特點類似[17]。實驗組rd11小鼠出生后8周,F-ERG檢查結果顯示視網膜外層殘留的細胞幾乎完全沒有感光功能;我們推測這些細胞并非來源于光感受器,而是其細胞核分布于ONL附近的Müller細胞、水平細胞等,因此得以長期保留。
本研究結果表明,LPCAT1缺陷主要影響視網膜外層光感受器的結構并引起視覺電信號的異常。視網膜內層的神經節細胞在正常情況下也存在少量LPCAT1表達,但目前尚不清楚神經節細胞在視網膜變性早期是否出現功能障礙,從而影響生物電信號向中樞傳遞。這一點對研究視網膜變性的發病機制及今后指導視網膜基因治療非常關鍵,將是我們下一步研究工作的重點。
溶血卵磷脂酰基轉移酶1(LPCAT1)是一種促進磷脂合成及重構的酶,能使溶血卵磷脂轉化成二棕櫚酰卵磷脂,在人類多處正常組織中存在表達[1, 2]。研究發現,小鼠Lpcat1基因自發突變可引起視網膜變性,小鼠13號染色體上Lpcat1基因第3外顯子由于單個核苷酸插入引起移碼突變是rd11小鼠視網膜致病原因[3, 4]。但目前有關LPCAT1的研究多集中于合成肺表面活性物質及腫瘤[2, 5-10]。而針對其與視網膜的關系研究還較為少見。為此,我們觀察Lpcat1基因自發突變小鼠視網膜組織和閃光視網膜電圖(F-ERG)改變情況,現將結果報道如下。
1 材料和方法
Lpcat1基因自發突變純合子的rd11新生小鼠60只,美國實驗動物中心提供,作為實驗組。與之同齡的野生型C57BL/6J小鼠60只,溫州醫科大學實驗動物中心提供,作為對照組。
小鼠8周齡時,每組取5只小鼠。采用CO2氣體處死小鼠并摘取眼球,置于4%多聚甲醛冰凍切片固定液中4 ℃過夜,將固定好的標本在30%蔗糖溶液中脫水5 h。眼球置于冷凍包埋液中-80 ℃包埋,冰凍切片機做厚度為10 μm連續切片,切片平行于視軸方向,貼片于0.05%多聚賴氨酸處理的載玻片上,室溫干燥1 h。冰凍切片置于0.3%Triton X-100穿透液中30 min增強細胞膜通透性,5%牛血清白蛋白工作液中37 ℃封閉2 h,滴加一抗兔抗小鼠LPCAT1抗體(美國Sigma公司),4 ℃孵育過夜,二抗紅色熒光染料Cy3標記的山羊抗兔IgG(德國Millipore公司)37 ℃孵育1 h,清洗后滴加4,6-聯脒-2-苯基吲哚(DAPI)染色細胞核。熒光顯微鏡下觀察視網膜內核層(INL)、內叢狀層(IPL)、外核層(ONL)、外叢狀層(OPL)、神經節細胞層(GCL)、光感受器內節(IS)及外節(OS)等組織,根據是否產生紅色熒光確定LPCAT1陽性或陰性表達。
出生后3、6、9 d和2、4、6、8周,每組各時間點取5只小鼠。采用CO2氣體處死小鼠并摘取眼球,置于含4%多聚甲醛、2%三氯乙酸、2%氯化鋅和20%異丙醇的石蠟切片固定液中,4 ℃固定24 h;將固定好的標本常規脫水、浸蠟和包埋,將眼球蠟塊平行于視軸方向切片,取經視網膜后極部,距視盤邊緣大約50 μm區域的切片,片厚4 μm,貼片于0.05%多聚賴氨酸處理的載玻片上。常規蘇木精-伊紅(HE)染色后中性樹膠封片,生物顯微鏡下觀察視網膜結構。
出生后2、4、6、8周,每組各時間點取5只小鼠,行單眼全視野F-ERG檢查。小鼠暗適應過夜,在暗紅光條件下用0.5%復方托吡卡胺滴眼液散瞳,分別以72 mg/kg和4 mg/kg的劑量腹腔注射氯胺酮和賽拉嗪混合液麻醉。小鼠全身麻醉后在右眼眼球角鞏膜緣放置由直徑2 mm細銀絲制成的記錄電極,雙耳正中頭皮下和尾部皮下分別放置自制針狀的參考和接地電極。補暗適應5 min后先記錄暗視桿體反應,然后明適應10 min后再記錄明視錐體反應。使用刺激器(Ganzfeld Q450,德國Roland公司),白色閃光的色溫為7000 K。參照文獻[11]的方法設置參數,誘導暗視桿體反應的光強度為0.01 cd·s/m2;明適應背景白光的亮度為30 cd/m2,標準閃光強度3.0 cd·s/m2誘導明視錐體反應;單次閃光刺激持續時間2 ms,刺激間隔10 s,重復5次后取平均值,通頻帶設為1~100 Hz。 選取反應較大的正向b波作為觀察指標。
采用 SPSS 13.0 統計軟件對數據進行統計分析,實驗數據均以均數±標準差(
2 結果
免疫組織化學染色發現,實驗組小鼠視網膜組織中LPCAT1呈陰性表達(圖 1);對照組小鼠視網膜組織中LPCAT1呈陽性表達,主要分布于IS,GCL可見少量表達(圖 2)。
圖1
實驗組小鼠出生后8周LPCAT1表達的熒光顯微鏡像。1A. 視網膜后極部和周邊部均呈陰性表達 免疫熒光染色 ×25;1B. 視網膜后極部距視盤邊緣大約50 μm區域殘存的INL、IPL、GCL各層均呈陰性表達 免疫熒光染色 ×200 ? ?圖 2 對照組小鼠出生后8周LPCAT1表達的熒光顯微鏡像。2A. 視網膜后極部和周邊部均呈陽性表達 免疫熒光染色 ×25;2B. 視網膜后極部距視盤邊緣大約50 μm區域IS可見高密度表達,GCL見較少量表達 免疫熒光染色 ×200
光學顯微鏡觀察發現,實驗組小鼠于出生后9 d左右視網膜ONL已基本形成(圖 3A~3C);出生后2周ONL有8~9層結構(圖 3D);出生后4周ONL減少至5~6層結構(圖 3E),光感受器內外節連接(IS/OS)厚度變窄,并可見一些細胞核殘骸向視網膜色素上皮(RPE)端靠近,OPL厚度也相應變窄;出生后6周ONL僅剩2層結構(圖 3F);出生后8周ONL殘留0~1層結構(圖 3G)。對照組小鼠視網膜組織中ONL也在出生后9 d左右形成(圖 4A~4C);出生后2~8周,ONL始終保持在11~12層結構(圖 4D~4G),且IS/OS及OPL厚度均基本正常。
圖3
實驗組小鼠視網膜組織結構光學顯微鏡像。3A~3G分別為出生后3、6、9 d和2、4、6、8周。出生后9 d左右視網膜ONL已基本形成,出生后4~8周ONL層數迅速減少,直至幾乎消失,IS/OS及OPL厚度均變窄 HE ×400
圖4
對照組小鼠視網膜組織結構光學顯微鏡像。4A~4G分別為出生后3、6、9 d和2、4、6、8周。出生后9 d左右視網膜ONL已基本形成,出生后2~8周時視網膜各層組織結構正常 HE ×400
全視野F-ERG檢測結果顯示,實驗組小鼠出生后2、4周暗視桿體系統反應存在,呈降低趨勢,出生后6~8周暗視桿體系統反應消失;對照組小鼠各時間點暗視桿體系統反應正常(圖 5)。 出生后2周,實驗組和對照組小鼠b波振幅分別為(72.8±15.6)、(105.2± 21.1) μV,實驗組較對照組降低,但差異無統計學意義(t=-2.760,P=0.025)。出生后4周,實驗組和對照組小鼠b波振幅分別為(20.6±6.4)、(231.8±32.0) μV,實驗組較對照組明顯降低,差異有統計學意義(t=-14.471,P=0.000)(圖 6)。
圖5
兩組小鼠暗適應狀態下全視野F-ERG檢測像。實驗組小鼠出生后2、4周桿體系統反應存在,但反應幅值較同齡對照降低,出生后6~8周桿體系統反應消失;對照組小鼠各時間點暗視桿體系統反應正常 ? ?圖 6 兩組小鼠暗視b波振幅比較 ? ?圖 7 兩組小鼠明適應狀態下全視野F-ERG檢測像。實驗組小鼠出生后2、4、6周錐體系統反應存在,但反應幅值隨年齡增長逐漸降低,出生后8周明視錐體系統反應消失;對照組小鼠各時間點明視錐體系統反應正常 ? ?圖 8 兩組小鼠明視b波振幅比較
全視野F-ERG檢測結果顯示,實驗組小鼠出生后2、4、6周明視錐體系統反應存在,呈降低趨勢,出生后8周明視錐體系統反應消失;對照組小鼠各時間點明視錐體系統反應正常(圖 7)。出生后2、4周,實驗組小鼠b波振幅分別為(46.8±7.2)、(78.0±8.2) μV;對照組小鼠b波振幅分別為(42.8±6.4)、(91.4±9.4) μV。兩組比較,差異均無統計學意義(t=0.930、-2.401,P=0.379、0.043)。出生后6周,實驗組和對照組小鼠b波振幅分別為(17.2±2.0)、(116.2±12.9) μV,實驗組較對照組明顯降低,差異有統計學意義(t=-17.008,P=0.000)(圖 8)。
3 討論
LPCAT1的相對分子質量為59×103,位于內質網,其功能為促進脂質合成和代謝,并影響Ca2+通道[12]。有研究發現,糖尿病小鼠視網膜的LPCAT1表達降低[13]。Friedman等[3]研究發現,Lpcat1基因突變與小鼠視網膜變性有關,他們用定量聚合酶鏈反應的方法在野生型小鼠視網膜組織的INL、ONL及GCL均檢測到Lpcat1基因表達。我們采用免疫熒光染色法檢測LPCAT1在全視網膜的表達情況,結果顯示實驗組rd11小鼠全視網膜組織LPCAT1呈陰性表達。而對照組小鼠視網膜組織中LPCAT1呈陽性表達,其中IS可見高密度表達,GCL較少表達,但在INL呈陰性表達,這可能與其表達量太低有關[3]。這一結果與LPCAT1位于內質網,而內質網在光感受器中的分布主要集中在IS的結果相吻合[12, 14]。
LPCAT1缺陷引起視網膜變性的詳細過程目前仍不太清楚。本研究選取小鼠出生后3、6、9 d和2、4、6、8周7個時間點,對視網膜組織顯微結構進行動態觀察。結果顯示,實驗組rd11小鼠視網膜光感受器細胞在出生后9 d左右形成,從時間上判斷與對照組野生型小鼠差別不大。說明LPCAT1缺陷并未引起視網膜組織發育的明顯滯后。從小鼠各年齡段視網膜組織形態分析,LPCAT1缺陷主要引起光感受器細胞快速喪失,這可能與細胞凋亡有關。已有研究發現,LPCAT1缺陷可導致Ca2+通道持續開放,細胞內Ca2+濃度過高;鈣超載后引起一系列生化反應,最終導致細胞凋亡通路的啟動[14, 15]。而由于LPCAT1在正常視網膜組織中的GCL較少表達,該蛋白缺陷對后者結構的影響可能不大,所以我們未發現GCL形態的明顯改變。
F-ERG是視網膜組織接受光刺激后產生的一組電位反應[16]。小鼠完全暗適應狀態下,用弱白光刺激,可以引出桿體系統反應;經過明適應,充分抑制桿體系統功能后,用標準閃光強度可以誘導出錐體系統反應[11]。本研究結果顯示,對照組野生型小鼠出生后2~6周,其視網膜功能尚未完全發育成熟,直至出生后8周F-ERG才基本穩定。我們發現,實驗組rd11小鼠視桿細胞變性始于出生后4周,視錐細胞變性始于出生后6周,在接下去不到2周的時間內視網膜功能幾乎完全喪失。而視網膜結構也顯示小鼠出生后4周光感受器減少最為顯著,這與在數量上占絕對優勢的視桿細胞的變性凋亡基本相符;出生后6~8周,視網膜殘留的具有椎體功能的1~2層光感受器也完全消失了。這些結果提示光感受器變性發生時間早,進展速度快。另外,視桿細胞變性要早于視錐細胞,這同臨床上原發性視網膜色素變性發病特點類似[17]。實驗組rd11小鼠出生后8周,F-ERG檢查結果顯示視網膜外層殘留的細胞幾乎完全沒有感光功能;我們推測這些細胞并非來源于光感受器,而是其細胞核分布于ONL附近的Müller細胞、水平細胞等,因此得以長期保留。
本研究結果表明,LPCAT1缺陷主要影響視網膜外層光感受器的結構并引起視覺電信號的異常。視網膜內層的神經節細胞在正常情況下也存在少量LPCAT1表達,但目前尚不清楚神經節細胞在視網膜變性早期是否出現功能障礙,從而影響生物電信號向中樞傳遞。這一點對研究視網膜變性的發病機制及今后指導視網膜基因治療非常關鍵,將是我們下一步研究工作的重點。